
Život na Zemi sa neustále vyvíja a rastliny predstavujú fascinujúci príklad prispôsobenia a evolučného pokroku. Vyššie rastliny, známe aj ako Cormobionta alebo Embryobionta, sa vyvinuli pred viac ako 500 miliónmi rokov zo zelených rias, konkrétne zo skupiny chary. Sú to prevažne autotrofné organizmy, v chloroplastoch ktorých sa nachádza dvojica chlorofylov a + b, kľúčových pre fotosyntézu. Počas evolúcie urobili vyššie rastliny kľúčový krok - opustili vodné prostredie a prispôsobili sa životu na suchej zemi, čo si vyžiadalo rozsiahle anatomické a fyziologické zmeny.
Na rozdiel od nediferencovanej stielky nižších rastlín, telo vyšších rastlín má zložitejšiu stavbu a nazýva sa kormus. Prvé suchozemské rastliny sa museli chrániť pred vysušením, čo vyriešili vznikom pokožky, ktorá tvorí ich vonkajší ochranný obal. Keďže rastlina už nebola celá ponorená vo vode, musela vybudovať efektívny systém rozvozu vody a v nej rozpustených látok po celom tele. Vytvorili sa tak diferencované cievne zväzky, známe ako vodivé pletivá, ktoré zabezpečujú transport. Telo vyšších rastlín sa skladá z diferencovaných buniek, ktoré vytvárajú špecializované pletivá a pravé orgány so špecifickými funkciami:
- Koreň - vyšpecializoval sa na príjem vodných roztokov z pôdy a na upevnenie rastliny k podkladu, čím jej poskytuje stabilitu.
- Stonka - prispôsobila sa na rozvoz živín medzi jednotlivými časťami rastliny a na primerané rozmiestnenie rastlinných orgánov (listov, kvetov) v priestore tak, aby optimalizovala prístup k svetlu.
- Listy - skoncentrovala sa v nich fotosyntéza, pričom ich rozšírenie do plochy ju urobilo oveľa efektívnejšou a umožnilo maximálnu absorpciu slnečného žiarenia.
Procesy spojené s vytváraním pohlavných buniek a rozmnožovaním sa taktiež situovali do špecifických orgánov. Výtrusné rastliny (Sporophyta) sú starobylou skupinou, ktorá vznikla asi pred 420 miliónmi rokov v silúre. Ako už názov napovedá, rozmnožujú sa pomocou jednobunkových výtrusov (spór), ktoré vznikajú vo výtrusniciach. Ich najväčšou evolučnou nevýhodou je pretrvávajúca závislosť od vodného prostredia pri oplodnení. Ich samčie pohlavné bunky (spermatozoidy) sú pohyblivé vďaka bičíkom a aby sa dostali k vajíčku, musia k nemu doslova doplávať vo vrstvičke vody, pričom stačí aj kvapka rosy alebo dažďa. Pre ich život je typické striedanie pohlavnej haploidnej generácie (gametofytu) s nepohlavnou diploidnou generáciou (sporofytom), jav, ktorý označujeme ako rodozmenu (metagenézu). Gametofyt u nich zvyčajne rastie nezávisle od sporofytu vo vonkajšom prostredí (exospóricky). Z viac ako 12 000 dnes známych druhov cievnatých výtrusných rastlín tvoria drvivú väčšinu (vyše 90 %) sladičorasty (paprade). Osobitnú kategóriu výtrusných rastlín tvoria machorasty, ktorým chýba rozsiahly vodivý systém tvorený xylémom a floémom, preto sa uvádzajú ako bezcievnaté výtrusné rastliny.
Semenné Rastliny: Kľúč k Nezávislosti od Vody
Semenné rastliny (Spermatophyta) dosiahli skutočný evolučný triumf, pretože sa pri oplodnení úplne oslobodili od vody. Ich samčie bunky (spermácie) už zväčša nemajú bičíky, čím sa eliminuje potreba vodného prostredia pre ich pohyb. Namiesto plávania vytvára rastlina peľovú trubicu, ktorá bunky bezpečne dopraví priamo k vajíčku. Tieto rastliny už neuvoľňujú spóry do prostredia; ich gametofyt je extrémne redukovaný a vyvíja sa chránený hlboko vo vnútri materskej rastliny (endospóricky).
Hlavným znakom semenných rastlín je to, že vytvárajú kvet (flos), v ktorom sa nachádzajú pohlavné orgány (tyčinky a piestiky) produkujúce peľ a vajíčka. Kvet predstavuje de facto jediný gametofytický útvar. Výsledkom rozmnožovania je mnohobunkové semeno, ktoré chráni embryo pred vyschnutím a obsahuje preň zásobu živín, čím zabezpečuje prežitie a počiatočný rast mladej rastlinky.
Nahosemenné rastliny (Gymnospermae) tvoria prechodný vývojový stupeň medzi papraďami a krytosemennými rastlinami. Dnes k nim patrí približne 760 druhov. Ich semená nevytvárajú oplodie, sú „nahé“ a ležia voľne odkryté na povrchu modifikovaných listov alebo semenných šupín, ktoré sú zvyčajne usporiadané do šišiek. Pri opeľovaní sa vo veľkej miere spoliehajú na vietor (anemogamia), čo je menej špecifické a efektívne. Vo vodivých pletivách dreva majú väčšinou len jednoduchšie cievice (tracheidy) a prebieha u nich len jednoduché oplodnenie, bez tvorby endospermu ako u krytosemenných rastlín. Väčšinu, asi 550 druhov (72 %), zaraďujeme do oddelenia borovicorastov (Pinophyta), pre ktoré je typické, že majú listy ihlicovité alebo šupinovité, najčastejšie vždyzelené.

Krytosemenné Rastliny: Vrchol Organizácie a Rozmanitosti Života
Ku krytosemenným rastlinám (Angiospermae, novšie Angiospermatophyta) patria evolučne najmladšie a najvyššie organizované rastliny, ktoré predstavujú najväčší taxón s približne 220 000 druhmi. Unikátnou charakteristikou tejto skupiny je, že vajíčka a následne aj semená nie sú voľne vystavené, ale sú ukryté v semenníku, z ktorého po dozretí vzniká plod (carpus). Plody, ktoré môžu byť suché alebo dužinaté, chránia semeno pred nepriaznivými vplyvmi a aktívne sa zúčastňujú na jeho rozširovaní, čím zabezpečujú efektívnu disperziu.
Krytosemenné rastliny vytvorili pestrofarebné a voňavé kvety s nektárom, vďaka čomu na opeľovanie využívajú živočíchy a hmyz (zoogamia). Táto stratégia je oveľa efektívnejšia a špecifickejšia ako opeľovanie vetrom, pretože umožňuje presnejší prenos peľu. Unikátom tejto skupiny je aj dvojité oplodnenie, proces, pri ktorom jedna spermatická bunka oplodní vajíčko (vzniká embryo) a druhá splynie s polárnymi jadrami, čím vzniká triploidné vyživovacie pletivo nazývané endosperm.
Moderné fylogenetické štúdie DNA, ako sú systémy Angiosperm Phylogeny Group (APG), napríklad APG II (2003), APG III (2009) a APG IV (2016), ukázali, že kým jednoklíčnolistové rastliny tvoria jednu ucelenú vývojovú vetvu (sú monofyletické), obrovská historická skupina dvojklíčnolistových rastlín pochádza z viacerých rôznych vývojových línií (sú polyfyletické). Najnovšie sa preto namiesto podtried používa pojem vetva. Taktiež sa upúšťa od termínu magnóliorasty (Magnoliophyta), ktorý nahrádza pojem krytosemenné rastliny (Angiospermatophyta). Systém APG rozdeľuje krytosemenné rastliny na niekoľko hlavných monofyletických vetiev:
- Bazálne krytosemenné rastliny - predstavujú evolučne najprimitívnejšie druhy s jednoduchšími cievicami a peľom len s jedným otvorom (napríklad lekno). Tvoria už len 0,5 % dnešných druhov krytosemenných rastlín.
- Magnoliidová vetva - zachovávajú si niektoré primitívne znaky (peľ s jedným otvorom), no vyznačujú sa tvorbou éterických olejov (napríklad magnólie, vavríny, čierne korenie). Tvoria asi 2,5 % druhov.
- Jednoklíčnolistové rastliny - táto skupina ostala zachovaná ako monofyletická vetva (napríklad trávy, obilniny, orchidey, cibule). Tvorí asi 28 % druhov.
- Pravé dvojklíčnolistové rastliny (eudikotyledony) - ich hlavným moderným identifikačným znakom je trikolpátny peľ (peľové zrno s tromi klíčiacimi otvormi). Táto obrovská skupina sa ďalej delí na menšie vetvy. Referencie pre tieto klasifikácie zahŕňajú práce ako Hrabovský M., Záhradníková E., Mičieta, K.: Proposal of Slovak Names for Angiosperm Phylogeny Group Classification of Flowering Plants: APG IV. (2016) a Cronquist, A: Some realignments in the dicotyledons. (1983).
Kvet Krytosemenných Rastlín: Architektúra Rozmnožovania
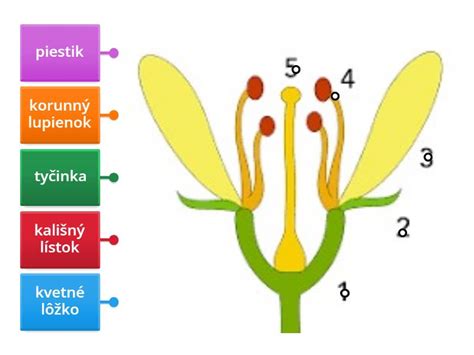
Kvet je rozmnožovacím orgánom listového pôvodu, ktorý vytvárajú hlavne krytosemenné rastliny. Nahosemenné rastliny nemajú typické kvety; ich vajíčka a neskôr semená sú uložené napríklad v šišticiach. Kvet krytosemenných rastlín sa skladá z týchto základných častí:
- Kvetná stopka - ňou sa kvet pripája na stonku, zabezpečuje prísun vody a živín.
- Kvetné lôžko - vzniklo zo stonky a nesie všetky ostatné časti kvetu, slúži ako opora.
- Kvetné obaly - vznikli premenou listov a chránia vnútorné časti kvetu. Môžu byť rôzne sfarbené a voňavé, aby prilákali opeľovače. Môžu byť rozlíšené na kalich a korunu.
- Kalich je väčšinou zelený, tvorí vonkajšie časti obalov. Môže byť trváci alebo prchavý; u niektorých rastlín sa môže premeniť napríklad na páper (púpava), ktorý pomáha pri rozširovaní semien.
- Koruna tvorí vnútorné obaly a môže byť pestro sfarbená (u hmyzoopelivých rastlín) alebo môže byť nenápadná, prípadne úplne chýba (u vetroopelivých rastlín). Na korunových lístkoch alebo v ich blízkosti sa môžu nachádzať nektáriá, ktoré produkujú sladký nektár, lákajúci opeľovače. Rozlíšený kvetný obal je typickým znakom dvojklíčnolistových rastlín. Jednoklíčnolistové rastliny môžu mať kvetný obal nerozlíšený, nevytvárajú kalich a korunu a nazývame ho okvetie (perigon), napríklad u tulipánov.
Samčie Rozmnožovacie Orgány: Tyčinky
Tyčinky sú samčie rozmnožovacie útvary, ktoré sa skladajú z nitky a peľnice. Nitka môže byť rôzne dlhá a u niektorých rastlín môže tyčinka vyčnievať von z kvetu, čo uľahčuje prenos peľu. V peľnici sa nachádzajú peľové komôrky, v ktorých sa vytvárajú peľové zrná. Súbor všetkých tyčiniek v kvete sa nazýva andreceum.
Samičie Rozmnožovacie Orgány: Piestik
Piestik vzniká u krytosemenných rastlín zrastením jedného alebo viacerých samičích plodolistov (megasporofyly). Nahosemenné rastliny piestik nemajú; ich plodolist je plochý (tvorí ho semenná šupina), na ňom sa nachádza nekryté vajíčko. Piestik sa skladá z:
- Blizny (a) - vrchná časť piestika, ktorá je často vlhká a lepkavá. Na blizne sa zachytávajú peľové zrná, čo je prvý krok k oplodneniu.
- Čnelky (b) - stredná časť piestika, ktorá je rôzne dlhá. Jej funkciou je dvíhať bliznu tak, aby sa na nej mohli zachytiť peľové zrná alebo aby sa blizna otrela o hmyz hľadajúci v kvete sladkú šťavu (nektár).
- Semenníka (c) - spodná, zväčšená časť piestika, v ktorej sú ukryté vajíčka. Usporiadanie semenníka vzhľadom na kvetné obaly a tyčinky môže byť:
- Vrchný - kvetné obaly a tyčinky vyrastajú pod semenníkom.
- Polospodný - kvetné obaly a tyčinky vyrastajú približne uprostred.
- Spodný - kvetné obaly a tyčinky vyrastajú nad semenníkom.
Vývoj Samičieho Gametofytu: Zložitá Cesta k Oosfére
V semenníku piestika sa vyvíja jedno alebo viac vajíčok. Vajíčko vzniklo zrastením výtrusníc. Prostredná výtrusnica tvorí vajíčkové jadro (nucellus), zatiaľ čo ostatné výtrusnice majú ochrannú funkciu a tvoria vajíčkové obaly - integumenty. Na vrchole vajíčka vytvárajú integumenty malý otvor, známy ako peľový vchod (micropyle), ktorý je kľúčový pre vstup peľovej trubice.
Vajíčkové jadro je vyplnené výtrusorodým pletivom, ktorého jedna bunka - materská bunka zárodočného mieška (megasporocyt) - sa redukčne delí (meióza). Vzniknú štyri haploidné bunky, z ktorých tri zaniknú a zostávajúca bunka tvorí jednobunkový zárodočný miešok s centrálnym jadrom. Tento proces sa nazýva megasporogenéza.
Následne v zárodočnom miešku prebehnú tri mitotické delenia, čím vznikne osem-jadrový sedem-bunkový útvar. V jeho strede vzniká diploidné centrálne jadro, ktoré je výsledkom splynutia dvoch polárnych jadier. K hornému koncu útvaru sa premiestni vajcová bunka - oosféra, ktorá je pripravená na oplodnenie, spolu s dvoma synergidami. Na opačnom póle zárodočného mieška sa vytvoria tri protistojné bunky (antipódy). V tomto štádiu vývinu ide o zrelý zárodočný miešok, ktorý je pripravený na oplodnenie. Samičí gametofyt krytosemenných rastlín je tak extrémne zredukovaný len na sedem buniek - zárodočný vak.

Vývoj Samčieho Gametofytu: Cesta Peľového Zrna
V peľniciach tyčiniek je diploidné peľotvorné pletivo, z ktorého sa po redukčnom delení (meióze) vyvíjajú štvorice (tetrády) peľových zrniek. Tento proces sa označuje ako mikrosporogenéza. V určitom štádiu vývinu peľníc sa z oblasti, ktoré sú v uhle peľnice, začne vyvíjať sporogénne pletivo. V každej skupine buniek môžeme nájsť vrstvu takzvaných podpokožkových iniciál - primárny archespór. Tieto bunky sa v priebehu vývinu delia a vznikajú dve vrstvy buniek: vnútorná vrstva buniek sa stáva primárnymi sporogénnymi bunkami (materské bunky peľových zŕn) a vonkajšia vrstva sa stáva primárnymi parietálnymi bunkami, ktoré tvoria steny peľnice a tapetum. Mitotickým delením primárnych sporogénnych buniek vznikajú mikrosporocyty (materské bunky peľových zŕn).
Mikrosporocyty sú navzájom prepojené plazmodezmami a rovnako sú prepojené s bunkami tapetu. Tesne pred meiózou sa obalia kalózovou vrstvou, pričom dochádza k prerušeniu plazmodeziem medzi mikrosporocytmi a zároveň medzi mikrosporocytmi a tapetom. Existuje však medzi nimi určité prepojenie pomocou cytomiktických kanálov, ktoré umožňujú výmenu informácií. Meiózou potom vzniká tetráda mikrospór. Mikrospóry ostávajú v tetrádach niekoľko hodín až dní, v závislosti od druhu rastliny, a následne dochádza k rozrušeniu kalózovej steny enzýmom kalázou, ktorý je produktom tapetu, a k ich uvoľneniu. Následne prebehne mitóza a vzniknú dve bunky.
Štruktúra a Funkcia Peľového Zrna
Jadrá peľových zrniek sa mitoticky delia predtým, ako zrnká opustia peľovú komôrku. Pri celularizácii dochádza k nerovnomernému rozdeleniu cytoplazmy medzi dve novovzniknuté bunky:
- Vegetatívna bunka - je väčšia a vypĺňa väčšiu časť objemu peľového zrna. Má sférické jadro a jadierko, málo organel. V procese dozrievania peľové zrno rastie, pričom sa zvyšuje metabolická aktivita, mení sa tvar jadra na laločnaté a zvyšuje sa počet organel. Prebiehajú biochemické procesy ako syntéza RNA a bielkovín. Pred ukončením rastu peľového zrna aktivita klesá, dochádza k dehydratácii a peľové zrno prechádza do dormantného stavu. Jej hlavnou funkciou je metabolické zabezpečenie rastu a vývinu peľového zrna a zabezpečenie klíčenia peľového zrna.
- Generatívna bunka - je menšia a predstavuje úzky pás cytoplazmy, oddelená od vegetatívnej bunky len cytoplazmatickou membránou. Je nutrične závislá od vegetatívnej bunky. Jej jadro je menšie, má vyšší obsah DNA a odlišné zloženie bielkovín. Funkciou generatívnej bunky je prenos genetickej informácie do samčích pohlavných buniek.
Peľové zrno, ktorého generatívna bunka je vo vnútri vegetatívnej bunky, sa nazýva dvojbunkový peľ. Ak prebehne ešte jedno mitotické delenie jadra generatívnej bunky, vzniknú dve spermatické bunky, a potom peľové zrno obsahuje vegetatívnu bunku a v nej uložené dve spermatické bunky; takýto peľ sa nazýva trojbunkový peľ. Samičí gametofyt krytosemenných rastlín je dokonca redukovaný len na 2-3 bunky (peľové zrnko).
Peľové zrno má vnútornú vrstvu nazývanú intima (pektocelulózová) a vonkajšiu vrstvu - exínu, ktorá obsahuje viac vrstiev. Exína nie je súvislá; miestami je povrch peľového zrna krytý len intimou, tieto miesta voláme apertúry. Cez apertúry peľové zrno klíči. Počet, tvar a rozloženie apertúr sú charakteristické pre dané taxonomické skupiny. Peľové zrná niektorých čeľadí môžu tvoriť zhluky - agregáty, napríklad diádach, tetrádach, polyádach, čo zvyšuje pravdepodobnosť opelenia. Veľké zhluky sa nazývajú massuli a ak je celý obsah peľového váčku zgrupovaný v jeden útvar, nazýva sa polynárium (napr. u orchidey).
Tapetum a jeho Význam
Tapetum je vnútorná výstelka peľnice, ktorej bunky sú parenchymatické, neskôr majú viskóznu cytoplazmu a doskovitý tvar. Jeho hlavnou funkciou je distribúcia živín a syntéza látok dôležitých pre vyvíjajúci sa peľ. Medzi jeho produkty patria kataláza (enzým, ktorý rozkladá kalózu), prekurzory exíny peľových zŕn a proteíny, ktoré sa stávajú súčasťou peľového zrna. Tapetum tiež syntetizuje „peľový tmel“, ktorý pozostáva z lipidovej a karoténovej zložky a ukladá sa na exínu peľových zŕn, pričom má význam pri prenose peľu u hmyzom opelivých rastlín. V procese vývinu peľu je úzko prepojený na procesy, ktoré sa realizujú pri vývine peľových zŕn. Pred ukončením diferenciácie peľových zŕn tapetum degeneruje a látky, ktoré obsahuje, sa využijú pri dostavbe peľu. Zvyšok sa zachová vo forme tryfínu, ktorý má úlohu pri interakcii medzi peľom, bliznou a čnelkou. Poruchy vo vývine tapetu môžu viesť k poruchám mikrosporogenézy a jedným z jej príčin je takzvaná samčia sterilita.

Opelenie: Kľúčový Krok k Oplodneniu
Opelenie (pollinatio) je prenesenie peľu z tyčinky na bliznu piestika. Ak je peľ prenesený z tyčinky na piestik toho istého kvetu, jedná sa o samoopelenie (autogamia). Ak je peľ prenesený na cudzí kvet, je to cudzoopelenie (allogamia). Rastliny sa samoopeleniu bránia rôznymi dômyselnými mechanizmami, ako je autoinkompatibilita (biochemický blok), časový nesúlad dozrievania orgánov (protogýnia, kde semenníky dozrievajú skôr než peľnice, alebo protandria, kde tyčinky dozrievajú skôr než semenníky) alebo priestorové usporiadanie kvetných orgánov (heterostýlia). Dômyselný priestorový mechanizmus na podporu cudzoopelenia využíva napríklad prvosienka jarná. Jednotlivé rastliny tvoria buď kvety s dlhými čnelkami a krátkymi tyčinkami, alebo presne naopak, čo priamo núti hmyz prenášať peľ výhradne medzi rôznymi jedincami. Hraničný prípad autogamie je kleistogamia, kedy k opeleniu dochádza ešte v nerozvitom kvetnom púčiku.
Opelenie môže prebiehať rôznymi spôsobmi:
- Živočíchmi (zoochória) - hmyzom, vtákmi alebo inými živočíchmi.
- Vetrom (anemochória) - je menej špecifické, ale efektívne u rastlín, ktoré produkujú veľké množstvo ľahkého peľu.
Ak na bliznu dopadne správny peľ, prítomný vápnik, bór a fytohormóny ho okamžite stimulujú ku klíčeniu. Peľové zrno po dopade na bliznu pohlcuje takzvanú polinačnú kvapku, ktorú vylučuje blizna, čo vedie k imbibícii. Polinačná kvapka obsahuje lipidické látky, mastné kyseliny, fenolické látky vo forme esterov a glykozidov. Aktivácia peľového zrna sa prejaví syntézou proteínov a RNA a zreteľným cirkulačným pohybom cytoplazmy. Nastáva fáza klíčenia, pri ktorej peľové zrnko klíči cez apertúru. Degradácia intimy prebieha mechanicky alebo enzymaticky (vplyvom enzýmov, pohybom cytoplazmy a zvýšením turgoru spôsobeným absorpciou polinačnej kvapky). Väčšinou k porušeniu intimy dochádza kombináciou mechanického a enzymatického pôsobenia. Enzým, ktorý naruší intimu, je sekrétom produkovaným bliznou.
Ak peľové zrno klíči cez jednu apertúru, ide o monosyfonické klíčenie; ak cez viac, polysyfonické klíčenie. Po vyklíčení peľového vrecúška sa do jeho puzdra presúva celý obsah peľového zrna. U dvojbunkového peľu dochádza k rozdeleniu generatívnej bunky na dve spermatické bunky, a toto peľové vrecúško potom predstavuje samčí gametofyt. Rýchlosť prerastania peľového vrecúška závisí od prísunu energetických a stimulačných látok. Rast peľového vrecúška je sústredený hlavne na apikálnu časť, kde sú koncentrované všetky fyziologicky aktívne látky - vitamíny, RNA, proteíny. Peľové vrecúško pri prerastaní pôsobí ako haustórium, to znamená, že sekrétmi rozrušuje bunky blizny aj čnelky. Ak peľové vrecúško prerastá cez dutú čnelku, ide o ektotrofné prerastanie; ak cez čnelku vyplnenú parenchýmom, ide o endotrofné prerastanie. Rozložené bunky absorbuje a využije pri ďalšom prerastaní. Piestik svojimi sekrétmi zabezpečuje chemotropické vedenie vrecúška k zárodočnému miešku. Z čnelky peľové vrecúško postupne prerastá medzi bunkami pletív semenníka a placenty k vajíčkam. Najčastejšie peľové vrecúško prerastie do zárodočného mieška cez mikropyl, čo sa nazýva porogamia. Ak prerastá cez integumenty, ide o mezogamiu, a ak prerastá cez chalazálnu oblasť vajíčka, je to chalazogamia. Mezogamia a chalazogamia predstavujú aporogamiu.
Dvojité Oplodnenie: Unikát Krytosemenných Rastlín
Dvojité oplodnenie je unikátny a zásadný proces, ktorý definuje rozmnožovanie krytosemenných rastlín. Tesne pred preniknutím peľového vrecúška do zárodočného mieška jedna zo synergíd degeneruje. Peľové vrecúško pokračuje v prerastaní cez filiformný aparát, pričom v tomto období má slizovitú konzistenciu a napomáha rastu peľového vrecúška. Peľové vrecúško vstupuje do degenerovanej synergidy, kde praská a uvoľní sa jeho obsah. Spermatické bunky prenikajú k plazmaléme vajcovej bunky a do centrálnej bunky; ostatné komponenty peľového vrecúška ostávajú v synergide. K oplodneniu dochádza buď bezprostredne po opelení (niekoľko minút/hodín) alebo až po niekoľkých dňoch až mesiacoch.
Dvojité oplodnenie má dve fázy:
- Syngamia: Ide o splynutie jadra samičej a samčej pohlavnej bunky. Jedna spermatická bunka splynie s oosférou (vajcovou bunkou), čím vznikne diploidná zygota. Z tejto zygoty sa následne vyvinie embryo, zárodok budúcej rastliny.
- Konfluácia: Druhá spermatická bunka oplodní diploidné centrálne jadro zárodočného mieška (ktoré je už pred oplodnením diploidné, vzniknuté splynutím dvoch polárnych jadier). Tým vznikne triploidná bunka, z ktorej sa utvorí triploidný sekundárny endosperm. Endosperm má kľúčovú funkciu, pretože vyživuje vyvíjajúce sa embryo.
Rozmnožovanie krytosemenných rastlín (kvitnúcich rastlín) AKTUALIZOVANÉ
Zygota a Vývoj Embrya: Počiatok Nového Života
Po syngamii, teda splynutí jednej spermatickej bunky s oosférou, vzniká diploidná zygota. Táto zygota predstavuje prvú bunku nového organizmu a je základom pre vývoj embrya. Zygota začne prechádzať sériou mitotických delení, pričom sa vyvíja mnohobunkový zárodok - embryo. Tento proces je starostlivo riadený a vedie k diferenciácii prvých orgánov rastliny.
Súčasne s vývojom embrya sa vyvíja aj endosperm z triploidnej centrálnej bunky po konfluácii. Endosperm je vitálne dôležité vyživovacie pletivo, ktoré poskytuje embryu potrebné živiny počas jeho raného vývoja a v priebehu klíčenia. Integumenty (vajíčkové obaly) sa diferencujú na obaly semena, nazývané osemenie. Z oplodneného vajíčka sa tak postupne vyvinie semeno, zatiaľ čo plodolisty sa transformujú na oplodie. Antipódy a synergidy, ktoré boli súčasťou zárodočného mieška, po oplodnení zaniknú, keďže ich funkcia je splnená.
Semeno a Plod: Ochrana a Výživa Embrya
Vajíčko sa ako celok po oplodnení premieňa na semeno. Semeno krytosemennej rastliny je mnohobunkový útvar, ktorý chráni embryo pred vyschnutím a obsahuje preň zásobu živín, čo mu umožňuje prežiť nepriaznivé podmienky a iniciovať rast, keď nastanú vhodné podmienky. Skladá sa z niekoľkých kľúčových častí:
- Osemenie (testa) - je pevná ochranná vrstva, ktorá vznikla z pôvodných vajíčkových obalov (integumentov). Chráni vnútorné časti semena pred mechanickým poškodením, vysychaním a patogénmi.
- Vyživovacie pletivo (endosperm) - je triploidné pletivo, ktoré vzniklo z centrálnej bunky zárodočného mieška po oplodnení druhou spermatickou bunkou. Slúži na výživu embrya počas jeho vývoja v semene a v počiatočných fázach klíčenia. V niektorých semenách sa endosperm absorbuje už počas vývoja embrya, a živiny sú uložené v klíčnych listoch embrya (napr. u fazule).
- Zárodok (embryo) - je základ budúcej rastliny, ktorý vznikol z diploidnej zygoty. Obsahuje prvotné orgány, ako sú radikula (základ koreňa), plumula (základ stonky a listov) a klíčne listy (kotyledóny), ktoré slúžia ako zásobárne živín alebo vykonávajú fotosyntézu po vyklíčení.
Pre úspešný vývin a následné klíčenie sa v semene, predovšetkým v endosperme, koncentrujú kľúčové živiny a veľké množstvo fosforu, ktoré sú nevyhnutné pre rast a metabolizmus mladej rastlinky. Semeno následne stratí väčšinu vody (jej obsah klesne na 5-20 %) a prechádza do ochranného stavu spánku, známeho ako dormancia. Tento stav umožňuje semenu prežiť nepriaznivé obdobia (sucho, chlad) a klíčiť len v optimálnych podmienkach. Výnimočne môže semeno vzniknúť aj úplne bez oplodnenia. V zoológii sa vývin z neoplodneného vajíčka nazýva partenogenéza, no v botanike tento jav produkcie klonálneho semena označujeme ako apomixia.
Plod (carpus) vzniká premenou piestika po oplodnení. Jeho hlavnou funkciou je výživa semien v čase ich dozrievania a uľahčenie ich rozširovania (chória). Plod sa vyvíja na nadzemných častiach rastlín, ale niektoré dozrievajú v zemi (geokarpia), napríklad u podzemnice olejnej (búrske oriešky). Stena plodu - oplodia (perikarp) sa člení na:
- Exokarp - vonkajšia vrstva, rôzne hrubá a sfarbená, často s ochrannou alebo lákavou funkciou.
- Mesokarp - stredná časť, často tvorená šťavnatým parenchýmom (u dužinatých plodov).
- Endokarp - vnútorná časť, ktorá môže byť tvorená:
- Blanitým jadrovníkom (malvice).
- Sklerenchymatickým endokarpom (kôstkovice), ktorý tvorí tvrdú kôstku.
- Parenchymatickým endokarpom (bobule).

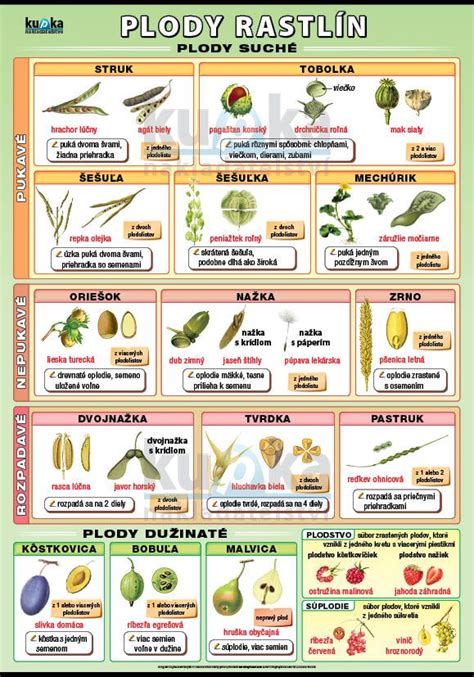
Rozdelenie Plodov
Plody sa delia na rôzne typy podľa ich štruktúry a premeny:
Dužinaté plody: Majú buď celé, alebo len vonkajšie a stredné oplodie dužinaté.
- Bobyľa - vzniká z jedno- alebo viacplodolistového piestika a má celé oplodie dužinaté (napr. vinič) alebo je vysychavé.
- Kôstkovica - je jedno- alebo viacplodolistový plod, má vonkajšie oplodie blanité, stredná časť je mäsitá, vnútorná časť vytvára kôstku, v ktorej je obyčajne jedno semeno (napr. slivka, čerešňa).
- Malvica - vzniká zo spodného alebo polospodného semenníka, ktorého stena zrastie s dužinatou kvetnou čiaškou (napr. jablko).
Podľa premeny:
- Pravé plody - vznikli premenou len piestika.
- Nepravé plody - vznikli premenou piestika a iných častí kvetu (napr. kvetného lôžka - jahoda). Nepravým plodom je aj malvica, v ktorej zrastá spodný semenník s bázou kvetných orgánov, a preto má pod dužinatou časťou pergamenovitý - tvrdý semenník spojený z 5 plodolistov.
Podľa perikarpu (stena plodu):
- Suché plody - s celistvým alebo čiastočne dužinatým oplodím.
- Pukavé - uvoľňujú semená v čase zrelosti.
- Šešuľa (kapustovité) - dvojplodolistový plod, rozdelený blanitou priehradkou (rámik), na ktorej sa nachádzajú semená.
- Šešuľka (kapustovité) - menšia ako šešuľa.
- Tobolka (mak, prvosienka) - vzniká zrastením viacerých plodolistov.
- Nepukavé - najčastejšie jednosemenné, v čase zrelosti semená neuvoľňujú, ale v celku sa oddeľujú od materskej rastliny.
- Nažka (astrovité) - jedno- alebo viacplodolistový plod s blanitým alebo kožovitým oplodím, u niektorých druhov s lietacím zariadením (púpava).
- Oriešok (lieska) - vytvorené sklerenchymatické oplodie.
- Zrno (lipnicovité) - oplodie tesne prilieha k osemeniu.
- Pukavé - uvoľňujú semená v čase zrelosti.
- Rozpadavé (poltivé) - sú jedno- alebo viacplodolistové, v čase zrelosti sa rozpadávajú na jednosemenné diely.
- Tvrdka (hluchavka)
- Pastruk (reďkev ohnica)
- Dvojnažka (javor, rasca)
- Zobákovitý plod (muškát)
Súplodie je súbor plodov vzniknutých z jedného súkvetia (napríklad strapec bobúľ hrozna, nažky v úbore slnečnice). Plodstvo je súbor plodov, ktoré vznikli z jedného kvetu s niekoľkými piestikmi (napr. plodstvo mechúrikov - záružlie, plodstvo kôstkovičiek - malina).
Klíčenie Semien a Rast Novej Rastliny
Klíčenie semien nastáva po prijatí vody (nabobtnanie), zosilnení dýchania a zvýšení enzymatickej aktivity v semene. Vonkajším prejavom je rast radikuly (korienka), ktorá sa ako prvá objavuje zo semena a upevňuje mladú rastlinku v pôde. Teplotné optimum klíčenia sa pohybuje medzi 25-28 °C pre rastliny mierneho pásma, avšak pre rôzne druhy sa môže líšiť.
Niektoré semená vyžadujú špecifické podmienky na klíčenie, ako je stratifikácia (dlhodobé vystavenie nízkym teplotám) alebo jarovizácia. Tieto procesy pomáhajú prekonať dormanciu semien, ktorá môže byť spôsobená tvrdým osemením alebo prítomnosťou inhibičných látok, ktoré bránia klíčeniu. Pred vybratím semien z dužinatých plodov by sa mali plody namočiť do vody, aby dužina zmäkla. Toto uľahčuje prijatie vody a urýchľuje klíčenie. Semená sa následne zmiešajú so substrátom a uložia na obdobie 4 až 12 týždňov, kým nezačnú klíčiť. Klíčidlo je zariadenie pre laboratórne stanovenie klíčivosti semien, ktorá vyjadruje podiel semien schopných vytvoriť morfologické znaky klíčenia v určenom čase.
Rodozmena a Genetická Variabilita u Krytosemenných Rastlín
Rodozmena (metagenéza) predstavuje životný cyklus charakterizovaný striedaním dvoch geneticky odlišných generácií: gametofytu a sporofytu. Fúziou haploidných gamét vzniká diploidná zygota, čím sa cyklus neustále opakuje a generácie sa striedajú. Gametofyt je pohlavná generácia, tvorená bunkami s jedným súborom chromozómov (haploidný počet, n). Vzniká mitotickým delením zo spóry. Neskôr vytvára pohlavné orgány (gametangiá), ktoré mitózou produkujú pohlavné bunky (gaméty): plemenníčky (anterídiá) tvoria samčie gaméty (často bičíkaté spermatozoidy vyžadujúce vodu) a zárodočníky (archegóniá) tvoria samičiu bunku (oosféra). Sporofyt je nepohlavná generácia, disponujúca homologickými chromozómami (diploidný počet, 2n). Vzniká mitotickým delením zygoty.
Prechod rastlín na súš znamenal evolučný tlak na ochranu citlivých fáz ich životného cyklu. Vývoj smeroval k posilneniu diploidného sporofytu a redukcii haploidného gametofytu. Prvé suchozemské rastliny, ako ryniorasty, mali ešte primitívnejšiu rovnakotvarú rodozmenu. Výraznou evolučnou výnimkou sú machorasty (Bryophyta), u ktorých dominuje haploidný gametofyt (zelený mach), pričom sporofyt (stopka s výtrusnicou) je fyziologicky úplne závislý od gametofytu. U cievnatých výtrusných rastlín už dominuje mohutný diploidný sporofyt, zatiaľ čo ich drobný gametofyt (prvorast) je síce zredukovaný, ale žije nezávislým životom.
Pri semenných rastlinách, vrátane krytosemenných, je gametofyt extrémne redukovaný a trvalo ukrytý v materskom sporofyte. Samičí gametofyt krytosemenných rastlín je zredukovaný len na sedem buniek (zárodočný vak) a samčí gametofyt dokonca len na dve až tri bunky (peľové zrnko). Pohlavné rozmnožovanie spája genetický materiál dvoch jedincov, čím zabezpečuje nevyhnutnú genetickú variabilitu. Nová rastlina vzniká zo zygoty, produkovanej splynutím dvoch haploidných gamét. Táto variabilita môže byť v záhradníctve niekedy nevýhodná, ak si želáme, aby si pestované rastliny uchovali určité vlastnosti. Jednotlivé rastliny rozmnožované semenami reagujú veľmi rozdielne a môžu byť nevyrovnané, so spravidla bujným rastom, rôzne dlhou juvenilnou fázou (neskorší vstup do rodivosti) a často aloríznym koreňovým systémom. Hospodárske vlastnosti takto získaných rastlín môžu byť horšie ako pri rodičovských rastlinách. Z tohto dôvodu sa generatívne rozmnožovanie pri ovocných rastlinách vzhľadom na niektoré jeho nevýhody využíva v súčasnosti v omnoho menšej miere.
Hybridy F1 vznikajú pri usmerňovanom krížení dvoch starostlivo udržiavaných geneticky čistých línií rovnakého druhu. Na získanie takéhoto osiva je potrebná komplexná šľachtiteľská technika, preto je omnoho drahšie. Vznikajú semená s vyššou životaschopnosťou než ich rodičia, často s prejavmi heterózy.
Vegetatívne Rozmnožovanie: Efektívna Alternatíva
Okrem pohlavného rozmnožovania existuje aj nepohlavné (vegetatívne) rozmnožovanie, ktoré vyžaduje len jeden materský organizmus. Bunkové delenia prebiehajú výlučne mitózou, vďaka čomu vznikajú geneticky identické klony. Je to energeticky výhodná stratégia na rýchlu kolonizáciu stabilného prostredia bez závislosti od opeľovačov. Nepohlavné rozmnožovanie je kľúčové pre druhy s poruchami meiózy, napríklad pri zmnožení chromozómových sád (polyploidia).
Jednoduchšie organizmy využívajú na množenie základné fyziologické formy: bunkové delenie (typické pre sinice a jednobunkové riasy), fragmentácia stielky (rozpad stielky na životaschopné časti, napr. sladkovodné spájavky) a výtrusy (jednobunkové útvary na šírenie, u nižších húb a organizmov tvorené aj mitoticky). Zložitejšiu stratégiu na nepohlavné šírenie majú niektoré machorasty.
Vyššie rastliny si na vegetatívne rozmnožovanie vyvinuli špecializované orgány, vzniknuté premenou stonky, koreňa či listov:
- Rozmnožovacia cibuľka - pozostáva z podcibulia a zdužnatých zásobných listov (napríklad tulipán, cesnak).
- Podzemok (rizóm) - vodorovne rastúca podzemná stonka. Z jej uzlov rastú korene a púčiky formujúce nové rastliny (napr. paprade, kostihoj, pýr).
- Podzemková hľuza - zhrubnutý zásobný podzemok stonkového pôvodu (napr. zemiak, kde „očká“ sú pazušné púčiky).
- Stonková hľuza - nadzemná alebo podzemná metamorfóza plniaca zásobnú aj reprodukčnú funkciu (napr. kaleráb, reďkovka ako koreňová hľuza).
- Poplaz (stolón) - horizontálna plazivá stonka, ktorá rastie po povrchu a zakoreňuje priamo vo svojich uzloch (napr. jahoda).
- Listy - na nepohlavné rozmnožovanie môžu slúžiť priamo aj listy (napr. kalanchoe).
V praxi sa veľmi často používa vegetatívne rozmnožovanie, pretože umožňuje získať rastliny, ktoré sú geneticky identické s rodičovskou rastlinou, čím sa zachovávajú požadované vlastnosti. Je často jednoduchšie a rýchlejšie, najmä pri rastlinách, ktoré sa ťažko rozmnožujú semenami.
Medzi bežné metódy patria:
- Delenie trsov - trs materskej rastliny sa rozdelí na niekoľko častí, každá s koreňmi a nadzemnou časťou.
- Potápanie - stonka je prikrytá pôdou, pričom je stále spojená s materskou rastlinou, aby vytvorila korene na nových výhonkoch.
- Odrezky - časti rastliny (koreňové, drevnaté, bylinné) sa režú a stimulujú k zakoreneniu. Pred výsadbou sa často použije rastový stimulátor.
- Štepenie - nepriame vegetatívne rozmnožovanie, pri ktorom sa spája časť vybranej odrody (očko, vrúbeľ) s vhodným podpníkom. Rôzne techniky zahŕňajú vrúbľovanie do boku, do rázštepu, na koziu nôžku, kopuláciu či plátkovanie. Očkovanie sa robí v období plnej miazgy. Dôležité je používať zdravé, vyzreté a kvalitné podpníky, očká a vrúble. Rez na vrúbľoch i očkách musí byť hladký a rezné plochy čisté, aby sa zabezpečilo krytie kambiálnych vrstiev pre úspešné zrastenie.
Semenárstvo a Ochrana Rastlín
Semenárstvo je odvetvie škôlkárstva, ktorého produktom sú semená. Osivo je rastlinný materiál určený na výsev a musí spĺňať prísne požiadavky na čistotu, klíčivosť a zdravotný stav. Osivo ovocných rastlín pre dopestovanie generatívnych podpníkov sa získava z plodov uznaných alebo registrovaných materských semenných stromov. Tieto stromy musia spĺňať prísne kritériá týkajúce sa pravosti druhu, odrody, rodivosti, odolnosti voči chorobám a škodcom, ako aj odolnosti voči nízkym teplotám a suchu. Okrem rozmnožovania je dôležitá aj starostlivosť o rastliny, ktorá zahŕňa ochranu pred chorobami a škodcami. Existujú rôzne príručky a systémy, ktoré pomáhajú identifikovať problémy s rastlinami a navrhnúť riešenia, ako aj správne techniky rezu, ktoré sú nevyhnutné pre ich zdravý rast a bohaté kvitnutie.
Rozmnožovanie krytosemenných rastlín (kvitnúcich rastlín) AKTUALIZOVANÉ
tags: #embryo #krytosemennych #rastlin