
Rozmnožovanie je základným pilierom života, procesom, ktorý umožňuje organizmom zachovať svoj druh a prenášať genetické informácie na potomstvo. Rastliny, rovnako ako živočíchy, vyvinuli rôznorodé a často komplexné mechanizmy na zabezpečenie svojej reprodukcie. Zatiaľ čo mnohé nižšie živočíchy sa spoliehajú na vegetatívne rozmnožovanie, u rastlín sa tento spôsob vyskytuje aj u vývojovo pokročilejších druhov. Nepohlavné rozmnožovanie, kde nový jedinec vzniká z časti tela rodičovskej rastliny prostredníctvom mitotického delenia buniek, zabezpečuje genetickú identitu potomstva. Tento proces je typický pre sinice a jednobunkové riasy, ktoré sa množia priamo delením.
Výtrusy, špecializované jednobunkové útvary určené na nepohlavné rozmnožovanie, vznikajúce v sporangiách, predstavujú ďalší významný mechanizmus šírenia u mnohých rastlinných skupín. Niektoré vyššie rastliny si dokonca vyvinuli špecializované útvary na vegetatívne rozmnožovanie, ako sú rozmnožovacie cibuľky (napr. cesnak, cibuľa), hľuzy (georgína, zemiak) alebo poplazy, na konci ktorých sa tvoria nové rastliny.
Na druhej strane, pohlavné rozmnožovanie prináša do reprodukčného procesu genetickú variabilitu. Nová rastlina sa vyvíja zo zygoty, ktorá vzniká splynutím dvoch pohlavných buniek - gamét. Samčia gaméta, nazývaná spermatozoid (u krytosemenných rastlín často súčasť peľového zrnka), sa spája so samičou gamétou, vajíčkovou bunkou. V procese meiózy, čiže redukčného delenia, sa počet chromozómov v gamétach zredukuje na polovicu (haploidný počet). Keďže nová rastlina dedí genetický materiál od dvoch rodičov, jej vlastnosti sa zvyčajne líšia od oboch rodičovských rastlín.
Pri pohlavnom rozmnožovaní u kvitnúcich rastlín hrá kľúčovú úlohu opelenie - prenesenie peľu z tyčinky na bliznu piestika. Môže ísť o samoopelenie (peľ z toho istého kvetu) alebo cudzoopelenie (peľ z iného kvetu tej istej alebo inej rastliny rovnakého druhu). Rastliny sa snažia zabrániť samoopeleniu rôznymi mechanizmami, napríklad nesúčasným dozrievaním samčích a samičích orgánov v tom istom kvete, alebo priestorovým usporiadaním tyčiniek a piestikov, ktoré podporuje prenos peľu medzi rôznymi jedincami prostredníctvom opeľovačov, ako je hmyz, vtáky či malé cicavce. Voda môže tiež slúžiť ako transportný prostriedok pre peľ u niektorých druhov.
Vývoj gametofytov: Od peľového zrna po zárodočný miešok
Proces dozrievania peľového zrna, ktoré predstavuje samčiu gametofytickú generáciu, začína v peľniciach po redukčnom delení. Vzniknuté haploidné jadrá sa ďalej mitoticky delia, pričom sa formujú vegetatívne (vyživovacie) a generatívne (rozmnožovacie) jadrá. Generatívne jadro sa môže ešte raz rozdeliť, čím vzniknú dve spermatické jadrá, ktoré sú pripravené na oplodnenie.

Dozrievanie vajíčka, ktoré je základom samičej gametofytickej generácie, prebieha odlišne u nahosemenných a krytosemenných rastlín. U nahosemenných rastlín sú plodolisty (megasporofyly) tvorené vretenom a šupinami, na ktorých sú uložené nekryté vajíčka. U krytosemenných rastlín plodolisty zrastajú do piestika. Vajíčko, mnohobunkový útvar obalený integumentmi s otvorom nazývaným mikropyla, obsahuje zárodočné pletivo (nucellus). Jedna bunka nucellu sa delí redukčne, čím vzniká haploidná megaspóra, ktorá sa ďalej vyvíja do zárodočného mieška. U väčšiny rastlín prežije len jedna megaspóra, zatiaľ čo ostatné zanikajú.
U nahosemenných rastlín sa jadro megaspóry mnohonásobne mitoticky delí, čím vzniká veľký počet haploidných jadier. Po vytvorení bunkových priehradok sa formuje primárny endosperm, ktorý slúži ako zásobný materiál. V endosperme sa následne diferencuje jeden alebo viac zárodočníkov (archegónium) s vajíčkovou bunkou (oosféra), čo predstavuje zrelý samičí gametofyt.
Naopak, u krytosemenných rastlín sa zostávajúca spóra trikrát mitoticky delí, čím vzniká ôsmich haploidných jadier, ktoré tvoria zrelý zárodočný miešok. Tento útvar je pripravený na oplodnenie po opelení.
Oplodnenie: Spojenie gamét a vznik nového života
Proces oplodnenia sa začína, keď zrelé peľové zrno dopadne na vlhkú bliznu a začne klíčiť. Vytvára sa peľové vrecúško, ktoré prerastá cez čnelku do semenníka a smeruje k vajíčku. Do peľového vrecúška sa presúvajú vegetatívne a obe spermatické jadrá. Po dosiahnutí vajíčka cez mikropylárny otvor, jedno spermatické jadro splynie s vajíčkovou bunkou, čím vzniká zygota.
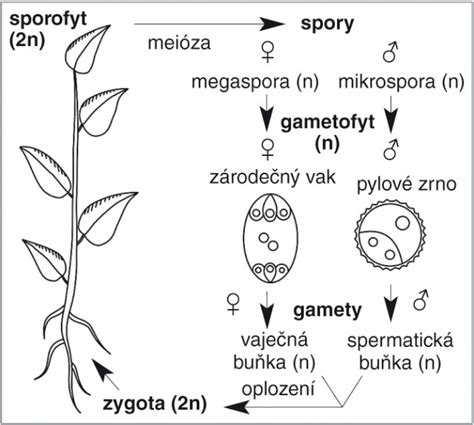
U nahosemenných rastlín splynie jedno spermatické jadro s oosférou a vytvorí sa zygota. Zásobné pletivo (endosperm) je u nich haploidné a vytvára sa pred oplodnením. U krytosemenných rastlín prebieha jedinečné dvojité oplodnenie. Jedno spermatické jadro sa spája s vajíčkovou bunkou a vytvára diploidnú zygótu (budúce embryo). Druhé spermatické jadro sa spája s centrálnym jadrom zárodočného mieška a vytvára triploidné (3n) jadrá, z ktorých sa vyvinie endosperm - výživné pletivo semena.
Evolučný vývoj a reprodukčné stratégie rastlinnej ríše
Historický vývoj rastlinnej ríše je úzko spätý s evolúciou ich reprodukčných stratégií. Ku koncu paleozoickej éry sa objavili prvé semenné rastliny, nahosemenné (Gymnospermy), ktoré začali konkurovať bezsemenným cievnatým rastlinám. Najmladšou a najrozšírenejšou skupinou sú krytosemenné rastliny (Magnoliophyta), charakteristické prítomnosťou kvetov, plodov a semien, ktoré začali dominovať od druhohôr. Tropické oblasti planéty sú domovom najväčšej rozmanitosti krytosemenných rastlín, zatiaľ čo ihličnaté rastliny (nahosemenné) sú v týchto lesoch zastúpené menej.
Reprodukčné cykly rastlín, podobne ako u iných živých organizmov, zahŕňajú narodenie, rast, rozmnožovanie a smrť. Pre zachovanie druhu si všetky živé organizmy vyvinuli mechanizmy na reprodukciu a udržanie svojho genetického zloženia, pričom rastliny si prispôsobili svoje reprodukčné stratégie environmentálnym podmienkam a evolučným tlakom.
Po úspešnom oplodnení u krytosemenných rastlín sa zygota vyvíja v embryo, ktoré je chránené a vyživované v rámci rastliny. Semeno, ktoré sa vyvinie z oplodneného vajíčka, je chránené plodom. Plody nielenže chránia semená, ale ich farba, chuť a vôňa lákajú živočíchy, ktoré ich šíria na nové miesta. Po dozretí a páde do úrodnej pôdy, alebo po prechode tráviacim systémom živočíchov, semená klíčia a začínajú nový reprodukčný cyklus.
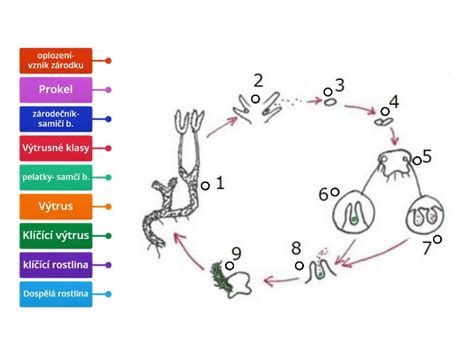
Asexuálna reprodukcia rastlín, ktorá neprináša genetické variácie, produkuje potomstvo geneticky totožné s materskou rastlinou. Tento spôsob je energeticky efektívny a umožňuje rýchlu kolonizáciu stabilného prostredia. U rastlín, ktoré netvoria semená, ako sú napríklad paprade a machy, sa nepohlavné rozmnožovanie uskutočňuje prostredníctvom spór. Tieto malé a ľahké útvary sú často šírené vetrom a sú schopné prežiť nepriaznivé podmienky až do optimálnych podmienok pre klíčenie.
Pri niektorých rastlinách, ako sú machy a paprade, je nevyhnutná prítomnosť vody pre transport samčích gamét, ktoré oplodnia samičie gaméty na inom gametofyte. Tieto gametofyty sú samy o sebe malé rastliny, ktorých hlavnou funkciou je umožniť reprodukciu.
Špecializované reprodukčné orgány a procesy u nahosemenných rastlín
Nahosemenné rastliny, ako napríklad borovica (Pinus silvestris), predstavujú dôležitý evolučný krok v reprodukcii rastlín. Na borovici, ktorá je sporofytom, vyrastajú samčie a samičie šištice (strobily). Samičie šištice, tvorené zdrevnateným vretenom so semennými šupinami, na ktorých sú uložené nekryté vajíčka, sa nachádzajú na vrchných konároch stromu. Vajíčka sú chránené jediným obalom (integumentom) s mikropylárnym otvorom.
Samčie šištice (mikrostrobily) sú menšie a tvorené krátkym vretenom s blanitými šupinami (mikrosporofylmi), ktoré nesú mikrosporangiá. Vznikajú v spodnejších častiach stromu.

Vývoj samičieho gametofytu u borovice začína vajíčkom, ktoré je krasinucelátne. Z jednej alebo viacerých subepidermálnych buniek nucelu sa vyvinie primárny archespór, ktorý sa delí, čím vzniká sekundárny archespór. Z neho sa niekoľko megasporocytov redukčne delí, čím vznikajú tetrády haploidných megaspór. U borovice sa vyvíja len bazálna megaspóra, ktorá mnohonásobným mitotickým delením vytvára voľnojadrové prothálium. Následne sa vytvoria bunkové steny, čím vzniká endosperm a archegóniá s oosférou.
Vývoj samčieho gametofytu začína v mikrosporangiách, kde sa z archespóru diferencuje tapetum a sporogénne pletivo. Redukčným delením vznikajú tetrády haploidných mikrospór. Tieto sa ďalej delia, pričom vznikajú vegetatívna bunka a generatívna bunka. Generatívna bunka sa delí na nástennú bunku a spermatogénnu bunku, z ktorej sa pri prerastaní peľového vrecúška vytvoria dve spermatické bunky. U niektorých primitívnejších nahosemenných rastlín sa samčie gaméty vyvíjajú ako obrvené spermatozoidy.
Opelenie u nahosemenných rastlín prebieha tak, že peľové zrná dopadajú medzi šupiny samičích šištíc a zachytávajú sa v polinačnej kvapke vylučovanej mikropylou. Pri vysýchaní kvapky sa peľové zrno dostane na povrch nucela a začne klíčiť, pričom vytvára peľové vrecúško. Spermatogénna bunka v peľovom vrecúšku sa rozdelí na dve spermatické bunky.
Oplodnenie nastáva, keď jedno spermatické jadro oplodní oosféru, čím vzniká zygota. Z oplodneného vajíčka sa vyvíja embryo a celé vajíčko sa premieňa na semeno. Z integumentu vzniká osemenie a z vonkajšej vrstvy sa diferencuje krídielko, ktoré napomáha šíreniu semien vetrom.
Proces opeľovania a oplodnenia 🌷| Postup krok za krokom
Rodozmena a jej evolučný význam
Rodozmena, čiže metagenéza, predstavuje životný cyklus rastlín charakterizovaný striedaním dvoch geneticky odlišných generácií: haploidného gametofytu (pohlavná generácia) a diploidného sporofytu (nepohlavná generácia). U prvých suchozemských rastlín bola rodozmena ešte primitívna, zatiaľ čo u machorastov dominuje haploidný gametofyt, pričom sporofyt je od neho závislý. U cievnatých výtrusných rastlín už dominuje diploidný sporofyt, zatiaľ čo gametofyt je zredukovaný, ale žije nezávisle. Pri semenných rastlinách je gametofyt extrémne zredukovaný a trvalo ukrytý v tele sporofytu. U krytosemenných rastlín je samičí gametofyt (zárodočný vak) zredukovaný na 7 buniek a samčí gametofyt (peľové zrnko) na 2-3 bunky.
Nepohlavné (vegetatívne) rozmnožovanie, ktoré nevyžaduje dva rodičovské organizmy a prebieha výlučne mitózou, produkuje geneticky identické klony. Je to energeticky výhodná stratégia na rýchlu kolonizáciu stabilného prostredia. Tento spôsob rozmnožovania je kľúčový pre druhy s poruchami meiózy, napríklad pri polyploidii.
Zložitejšie stratégie nepohlavného rozmnožovania zahŕňajú špecializované orgány, ako sú rozmnožovacie cibuľky, podzemky, hľuzy či poplazy. Tieto štruktúry, tvorené premenou stoniek, koreňov alebo listov, umožňujú rastlinám efektívne sa šíriť a kolonizovať nové územia. Aj listy môžu priamo slúžiť na nepohlavné rozmnožovanie.
Pohlavné rozmnožovanie, spájajúce genetický materiál dvoch jedincov, zabezpečuje nevyhnutnú genetickú variabilitu. Proces opelenia, samoopelenie alebo cudzoopelenie, je kľúčový pre úspešné oplodnenie a následný vývoj semena. Unikátny proces dvojitého oplodnenia u krytosemenných rastlín, kde jedno spermické jadro oplodní vajíčkovú bunku a druhé centrálne jadro zárodočného mieška, vedie k vzniku embrya a výživného endospermu. Z oplodneného vajíčka sa vyvinie semeno chránené osemením, ktoré po dozretí a strate vody prechádza do štádia dormancie, pripravené na klíčenie pri vhodných podmienkach. Výnimočne môže semeno vzniknúť aj bez oplodnenia prostredníctvom apomixie.
tags: #opelenie #a #oplodnenie #nahosemennych #rastlin