
Rozmnožovanie rastlín je základný a komplexný proces, ktorým vznikajú nové rastlinné jedince z materskej rastliny. Tento proces je kľúčový pre zachovanie druhu a prenos dedičných vlastností z rodičov na potomstvo, zabezpečujúc kontinuitu druhov. Môže prebiehať dvoma hlavnými spôsobmi: generatívne (pohlavné) alebo vegetatívne (nepohlavné). Kým nepohlavné rozmnožovanie vytvára geneticky identické klony, pohlavné rozmnožovanie prináša genetickú variabilitu, ktorá je nevyhnutná pre adaptáciu druhov na meniace sa podmienky prostredia.
I. Oplodnenie: Kľúč k Novému Životu
Pohlavné rozmnožovanie zahŕňa vznik novej rastliny zo zygoty, ktorá vzniká splynutím dvoch gamét - samčej a samičej pohlavnej bunky. Kľúčovou súčasťou tohto procesu je kvet, ktorý je špecializovaným orgánom pre rozmnožovanie. V kvetoch sa nachádzajú samčie orgány nazývané tyčinky, ktoré produkujú peľ, a samičí orgán nazývaný piestik. Prenos peľu z tyčinky na piestik sa nazýva opelenie, a nasleduje po ňom oplodnenie - splynutie samčej a samičej pohlavnej bunky. Samčia pohlavná bunka u rastlín sa nazýva oosféra, zatiaľ čo samčie bunky sú spermatozoidy alebo spermie, obsiahnuté v peľových zrnách.
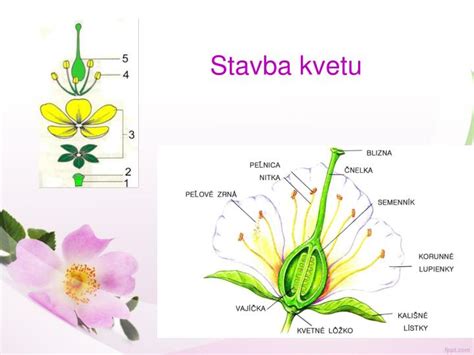
Vývoj Pohlavných Gamét
Aby mohlo dôjsť k oplodneniu, musia pohlavné bunky najprv dozrieť. Tento proces prebieha v špecializovaných štruktúrach rastliny.
Dozrievanie Peľového Zrna (Vznik Samčieho Gametofytu)
Peľové zrná vznikajú v peľnici po procese redukčného delenia (meiózy). Vzniknuté bunky majú haploidné jadro, ktoré sa následne mitoticky rozdelí na dve haploidné jadrá: vegetatívne (vyživovacie) a generatívne (rozmnožovacie). Generatívne jadro sa ešte raz rozdelí a vytvoria sa dve tzv. spermatické jadrá, ktoré slúžia ako samčie pohlavné bunky (gaméty) pre oplodnenie. Zrelé peľové zrnko krytosemenných rastlín tvoria tri bunky: jedna vegetatívna bunka (zabezpečuje rast peľovej trubice) a dve samčie neobrvené bunky, nazývané spermácie. Povrch peľových zŕn chráni pevná vonkajšia exina a vnútorná tenká intina.

Dozrievanie Vajíčka (Vznik Samičieho Gametofytu)
U semenných rastlín vznikajú vajíčka na plodolistoch. U nahosemenných rastlín sú plodolisty tvorené vretenom so šupinami, na ktorých sú uložené nekryté vajíčka. U krytosemenných rastlín plodolisty zrastajú a tvoria piestik (gynaeceum), pričom sa na každom plodoliste nachádza zárodočné pletivo (placenta), kde sa vytvárajú vajíčka.
Vajíčko je mnohobunkový útvar obklopený obalmi - integumentmi. V obaloch je otvor, mikropyla (peľový vchod), cez ktorý môžu samčie gaméty preniknúť do vajíčka. Vnútri vajíčka sa nachádza pletivo nucelus, ktorého jedna bunka (materská bunka) sa redukčne delí, čím vzniká haploidná megaspóra - nezrelý zárodočný miešok. U väčšiny rastlín prežije len jedna megaspóra, ktorá sa ďalej vyvíja. U krytosemenných rastlín sa zostávajúca spóra mitoticky delí trikrát, čím vzniká útvar s ôsmimi haploidnými jadrami, známy ako zrelý zárodočný miešok. Tento útvar, po opelení, je pripravený na oplodnenie. Samičí gametofyt krytosemenných rastlín je zredukovaný len na 7 buniek (zárodočný vak). U nahosemenných rastlín sa jadro zostávajúcej spóry mnohonásobne mitoticky delí, čím vzniká veľký počet haploidných jadier. Po vytvorení bunkových priehradok vzniká zásobné pletivo - primárny endosperm. V ňom sa postupne diferencuje jeden alebo viac zárodočníkov (archegónium) s vajcovou bunkou (oosféra). Tento útvar je zrelý zárodočný miešok u nahosemenných rastlín. Archegónium je samičí pohlavný orgán rastlín, zatiaľ čo oosféra je samičia gaméta.

Opelenie: Prenos Životodarnej Sily
Opelenie (pollinatio) je základným krokom pri pohlavnom rozmnožovaní kvitnúcich rastlín. Predstavuje prenesenie peľu z tyčinky na bliznu piestika. U nahosemenných rastlín sa peľové zrnko zachytáva priamo na vajíčku.
Typy Opelenia: Samoopelenie a Cudzoopelenie
Existujú dva hlavné typy opelenia:
- Samoopelenie (Autogamia): Pri samoopelení sa peľ prenáša z tyčinky na piestik toho istého kvetu. Špecifickým prípadom samoopelenia je kryptogamia (skrytoopelivosť), pri ktorej dochádza k opeleniu ešte v púčiku, pred rozkvitnutím kvetu.
- Cudzoopelenie (Allogamia): Cudzoopelenie nastáva, ak je peľ prenesený na piestik cudzieho kvetu, či už na inej rastline toho istého druhu, alebo na rastline iného jedinca.
Mechanizmy Prenosu Peľu
Peľ môže byť prenášaný rôznymi spôsobmi, ktoré rastliny vyvinuli ako adaptáciu na svoje prostredie:
- Vetrom (vetroopelivé rastliny): Tieto rastliny produkujú obrovské množstvo ľahkého peľu, ktorý je ľahko unášaný vzdušnými prúdmi. Typickým príkladom sú trávy.
- Živočíchmi: Ide o najrozmanitejší spôsob prenosu, pričom opeľovačmi môžu byť hmyz (ako včely, motýle, chrobáky), vtáky (napr. kolibríky) alebo dokonca malé cicavce. Rastliny lákajú opeľovačov farbou, vôňou a nektárom.
- Vodou: U niektorých vodných rastlín môže peľ preniesť voda.
Prevencia Samoopelenia
Samoopelenie je nežiaduce, pretože obmedzuje genetickú variabilitu, ktorá je pre druh dôležitá z hľadiska dlhodobej adaptácie. Preto sa rastliny fylogeneticky prispôsobili tak, aby sa mu čo najviac zabraňovalo. Medzi mechanizmy patrí:
- Autoinkompatibilita: Ide o biochemický blok peľu, kedy rastlina rozpozná svoj vlastný peľ a zabráni jeho klíčeniu alebo rastu peľovej trubice.
- Časový nesúlad dozrievania: V tom istom kvete nemusia tyčinky a piestik dozrievať naraz (napríklad u jabloní). To zabezpečí, že peľ z daného kvetu nie je schopný oplodniť vlastnú bliznu v čase jej receptivity.
- Priestorový mechanizmus (herkogamia): Tyčinky a piestiky sa nachádzajú v rôznych vzdialenostiach, čo podporuje prenos peľu medzi rôznymi jedincami. Dômyselný priestorový mechanizmus na podporu cudzoopelenia využíva napríklad prvosienka jarná. Jednotlivé rastliny tvoria buď kvety s dlhými čnelkami a krátkymi tyčinkami, alebo presne naopak, čo priamo núti hmyz prenášať peľ výhradne medzi rôznymi jedincami.
Oplodnenie a Vývoj Novej Rastliny
Ak na bliznu dopadne správny peľ, peľové zrnko vyklíči a vytvorí peľovú trubicu. Prítomný vápnik, bór a fytohormóny ho okamžite stimulujú ku klíčeniu. Peľové vrecúško prerastá cez čnelku až do semenníka. Do peľového vrecúška sa presúvajú vegetatívne jadro a obe už vytvorené spermatické jadrá. Peľové vrecúško prerastá až k obalom vajíčka, praská a spermatické jadrá sa dostávajú cez mikropylárny otvor dovnútra do vajíčka.
Dvojité Oplodnenie u Krytosemenných Rastlín
Proces oplodnenia u krytosemenných rastlín je unikátny a nazýva sa dvojité oplodnenie, pričom prebiehajú dve splynutia súčasne:
- Jedno spermatické jadro splynie s vajíčkovou bunkou (oosférou), čím vzniká diploidná zygota, z ktorej sa vyvinie embryo.
- Druhé spermatické jadro splynie s centrálnym jadrom zárodočného mieška a vytvorí sa triploidné (3n) endosperm, ktorý slúži ako výživné pletivo pre embryo.
Animácia dvojitého oplodnenia
Oplodnenie u Nahosemenných Rastlín
U nahosemenných rastlín dochádza k jednoduchému oplodneniu, kedy jedno spermatické jadro splynie s vajcovou bunkou za vzniku zygoty. Zárodočné pletivo (endosperm) je haploidné a vzniká pred oplodnením, na rozdiel od krytosemenných rastlín.
Vývin Semena a Dormancia
Z oplodneného vajíčka sa vyvíja semeno, chránené pevným osemením (testa), ktoré vzniklo z pôvodných vajíčkových obalov. Pre úspešný vývin a následné klíčenie sa v semene, predovšetkým v endosperme, koncentrujú kľúčové živiny a veľké množstvo fosforu. Semeno následne stráca väčšinu vody (jej obsah klesne na 5-20 %) a prechádza do ochranného stavu spánku (dormancie). Tento stav umožňuje semenám prežiť nepriaznivé podmienky a vyklíčiť až vtedy, keď sú podmienky optimálne.
Apomixia ako Alternatívna Cesta
Výnimočne môže semeno vzniknúť aj úplne bez oplodnenia. V zoológii sa vývin z neoplodneného vajíčka nazýva partenogenéza, no v botanike tento jav produkcie klonálneho semena označujeme ako apomixia. Príkladom rastlín schopných apomixie sú jastrabník a alchemilka. Semená môžu vznikať aj z buniek zárodočného mieška, čo je ďalšia forma apomixie.
II. Životný Cyklus (Ontogenéza) Rastliny
Zmeny, ktoré sa uskutočňujú v rastline od jej vzniku po jej zánik, označujeme ako ontogenetický vývin alebo ontogenéza. Ontogenéza je typická rastom a vývinom. Rast predstavuje pribúdanie hmoty rastliny. Tieto zmeny majú kvantitatívny charakter, lebo pribúda počet buniek a bunky sa rastom zväčšujú. Vývin zase predstavuje zmeny kvalitatívne, ako je diferenciácia buniek a orgánov. Všetky bunky rastliny, ktoré sa podieľajú na stavbe rastlinného tela, majú vlastnosť totipotencie, čo znamená, že v jadre obsahujú kompletnú genetickú informáciu a za určitých podmienok môžu z nich vzniknúť kompletné rastliny.
Cyklus začína semenom, ktoré po klíčení vytvára mladú rastlinu schopnú fotosyntetizovať. Zygota, ktorá vznikla oplodnením, sa hneď začína deliť. V priebehu embryonálneho vývinu sa postupne vyvíja celá klíčna rastlina, ktorá vyklíči zo semienka. Následne prebieha postembryonálny vývin, pri ktorom sa bunky v meristémoch stonky ďalej delia a špecializujú na vykonávanie určitých funkcií (diferenciácia). Neskôr rastlina začína kvitnúť, aby zabezpečila zachovanie svojho druhu prostredníctvom pohlavného rozmnožovania, prípadne sa rozmnožuje vegetatívne.
III. Širší Kontext Rozmnožovania Rastlín
Rozmnožovanie je jednou z hlavných podmienok existencie a zachovania druhu. Rastliny si počas svojej evolúcie vyvinuli rozmanité stratégie, aby prežili a kolonizovali rôzne prostredia.
Pohlavné (Generatívne) Rozmnožovanie Rastlín
Pohlavné rozmnožovanie je proces, pri ktorom nové jedince vznikajú splynutím dvoch pohlavných buniek (gamét). Vďaka tomu vzniká dcérsky organizmus, ktorý zvyčajne nesie kombináciu genetického materiálu dvoch rodičovských organizmov, čo vedie k genetickej variabilite. V širšom zmysle zahŕňa termín pohlavné rozmnožovanie každý typ rozmnožovania, pri ktorom sa nový jedinec vyvíja z gamét.
Genetické Princípy a Úloha Meiózy
Vlastnosti každého živého organizmu sú určené jeho deoxyribonukleovou kyselinou (DNA). Určité úseky DNA sa nazývajú gény. DNA v jadre eukaryotických organizmov je rozdelená do samostatných, rôzne dlhých úsekov, ktoré sa v spojení s bielkovinami formujú do hmoty nazývanej chromatín. V čase delenia bunky sa chromatín mení na zreteľne odlíšiteľné útvary - chromozómy. Počet chromozómov je u rôznych organizmov rôzny.
Pohlavné bunky, gaméty, vznikajú špeciálnym redukčným delením nazývaným meióza. Pri meióze sa počet chromozómov v bunke znižuje na polovicu, čím gaméty získavajú haploidnú sadu chromozómov (n). Splynutím dvoch haploidných gamét (oplodnenie) vzniká zygota s diploidnou sadou chromozómov (2n), z ktorej sa vyvíja nový jedinec. Pri meióze namiesto dvoch dcérskych buniek vznikajú až štyri dcérske bunky, každá len s haploidným počtom chromozómov. Existujú aj prípady, pri ktorých pohlavné bunky nevznikajú meiózou, alebo sú gaméty rovnakej veľkosti a tvaru (izogamia). Spojenie je možné len medzi dvoma bunkami rozdielneho pohlavia. Ak je jedinec schopný tvoriť samčie aj samičie pohlavné bunky, hovoríme o obojpohlavnosti (hermafrodizme).
Rodozmena (Striedanie Generácií)
U rastlín je typické striedanie pohlavnej a nepohlavnej generácie, nazývané rodozmena (metagenéza). Rodozmena predstavuje životný cyklus charakterizovaný striedaním dvoch geneticky odlišných generácií:
- Gametofyt (pohlavná generácia): Je to haploidná generácia (n), ktorá vzniká mitotickým delením zo spóry. Vytvára pohlavné orgány (gametangiá), ktoré mitózou produkujú pohlavné bunky (gaméty): plemenníčky (anterídiá) tvoria samčie gaméty (spermatozoidy) a zárodočníky (archegóniá) tvoria samičiu bunku (oosféru).
- Sporofyt (nepohlavná generácia): Je to diploidná generácia (2n), ktorá vzniká mitotickým delením zygoty. Sporofyt disponuje homologickými chromozómami.
Fúziou haploidných gamét vzniká diploidná zygota, čím sa cyklus neustále opakuje a generácie sa striedajú.

Evolúcia Rodozmeny
Prechod rastlín na súš znamenal evolučný tlak na ochranu citlivých fáz a viedol k posilneniu diploidného sporofytu a redukcii haploidného gametofytu. Prvé suchozemské rastliny, ako ryniorasty, mali ešte primitívnejšiu rovnakotvarú rodozmenu. Výraznou evolučnou výnimkou sú machorasty (Bryophyta), u ktorých dominuje haploidný gametofyt (zelený mach). Sporofyt (stopka s výtrusnicou) je fyziologicky úplne závislý od gametofytu. U cievnatých výtrusných rastlín už dominuje mohutný diploidný sporofyt. Ich drobný gametofyt (prvorast) je síce zredukovaný, ale žije nezávislým životom. Pri semenných rastlinách je gametofyt extrémne zredukovaný a trvalo ukrytý v materskom sporofyte. Samičí gametofyt krytosemenných rastlín je zredukovaný len na 7 buniek (zárodočný vak) a samčí gametofyt dokonca len na 2-3 bunky (peľové zrnko).
Nepohlavné (Vegetatívne) Rozmnožovanie Rastlín
Nepohlavné rozmnožovanie je proces, pri ktorom potomstvo vzniká z telových buniek rodičovskej rastliny. Je charakteristické tým, že nový jedinec je geneticky zhodný s materskou rastlinou, pretože bunkové delenia prebiehajú výlučne mitózou. Pri nepohlavnom rozmnožovaní vzniká nová rastlina z časti tela materskej rastliny čiže z buniek, ktoré vznikli mitotickým delením a majú rovnakú genetickú informáciu.
Princípy a Charakteristika Nepohlavného Rozmnožovania
Nepohlavné rozmnožovanie vyžaduje len jeden materský organizmus a je považované za energeticky výhodnú stratégiu na rýchlu kolonizáciu stabilného prostredia bez závislosti od opeľovačov. Je tiež kľúčové pre druhy s poruchami meiózy, napríklad pri zmnožení chromozómových sád (polyploidia). Na rozdiel od pohlavného spôsobu reprodukcie, ktoré je umožnené až po dosiahnutí pohlavnej dospelosti, nepohlavné rozmnožovanie môže prebiehať už od štádia zygoty.
Formy Nepohlavného Rozmnožovania u Jednoduchších Organizmov
Jednoduchšie organizmy využívajú na množenie základné fyziologické formy:
- Bunkové delenie: Typické pre sinice a jednobunkové riasy.
- Fragmentácia stielky: Rozpad stielky na životaschopné časti. Využívajú ju napríklad mnohobunkové sladkovodné spájavky, ktorých vlákno po roztrhnutí jednoducho dorastie na novú riasu.
- Výtrusy: Jednobunkové útvary na šírenie. Vznikajú vo výtrusniciach a u nižších húb a organizmov môžu byť tvorené aj mitoticky. Nekvitnúce rastliny, ako machy, paprade a prasličky, sa rozmnožujú výtrusmi. Nový jedinec môže vznikať zo spóry (výtrusu), ktorá sa tvorí v sporangiofóroch (výtrusniciach). Zložitejšiu stratégiu na nepohlavné šírenie majú aj niektoré machorasty.
Špecializované Vegetatívne Orgány Vyšších Rastlín
Vyššie rastliny si na vegetatívne rozmnožovanie vyvinuli špecializované orgány, ktoré vznikli premenou stonky, koreňa či listov:
- Rozmnožovacia cibuľka: Pozostáva z podcibulia a zdužnatých zásobných listov. Tento typ množenia je charakteristický pre čeľaď ľaliovité (napríklad obľúbený tulipán alebo cesnak, ktorý vytvára zložené dcérske cibuľky) a skorú jarnú čeľaď amarylkovité (snežienka, narcis).
- Podzemok (rizóm): Je vodorovne rastúca podzemná stonka. Z jej uzlov rastú korene a púčiky formujúce nové rastliny. Je to bežný jav predovšetkým u papradí, z kvitnúcich rastlín ho využíva napríklad liečivý kostihoj alebo mimoriadne odolný burinný pýr.
- Podzemková hľuza: Je zhrubnutý zásobný podzemok stonkového pôvodu. Najlepším príkladom je zemiak, ktorého známe „očká“ sú v skutočnosti pazušné púčiky novej rastliny. Podzemkové hľuzy tvorí aj georgína.
- Stonková hľuza: Je nadzemná alebo podzemná metamorfóza plniaca zásobnú aj reprodukčnú funkciu. Typickým zástupcom s nadzemnou hľuzou je kaleráb. Veľmi podobný mechanizmus, avšak vo forme koreňovej hľuzy, si vytvorila reďkovka.
- Poplaz (stolón): Ide o horizontálnu plazivú stonku, ktorá rastie po povrchu a zakoreňuje priamo vo svojich uzloch. Tento efektívny mechanizmus plošného šírenia má napríklad jahoda.
- Listy: Na nepohlavné rozmnožovanie môžu slúžiť priamo aj listy, ako je to u africkej fialky (senpólie), kde z jedného listu možno získať mnoho nových rastlín.
- Odrezky: Rozmnožovanie časťami tela rastliny, ako sú koreňové odrezky (chren), stonkové odrezky (muškát, okrasné dreviny) alebo listové odrezky (izbové rastliny ako senpólia). Odrezky sa odoberajú zo stredu rozkonáreného vrcholu zdravých rastlín a po zakorenení (vo vode alebo vlhkom substráte) tvoria nové jedince. Na urýchlenie zakoreňovania sa používajú stimulátory.
- Delenie trsov: Bežné u okrasných tráv.
- Odnože: Vznikajú z hlavného výhonku materskej rastliny a často už majú vlastné korene a listy. Oddeľujú sa a samostatne vysádzajú.
- Potápanie: Jednoročné výhonky sa ohýbajú do pôdy, kde zakorenia. Po oddelení od materskej rastliny sa nové kríky presadia. Používa sa napríklad pri ríbezliach alebo viniči.
- Štepenie a očkovanie: Techniky nepriameho vegetatívneho rozmnožovania, pri ktorých sa spája časť ušľachtilej odrody (púčik alebo vrúbeľ) s vhodným podpníkom. Vhodné pre bonsaje, palmy a ovocné dreviny.
Moderné Metódy: Rozmnožovanie in vitro (Mikropropagácia)
V súčasnosti sa pre intenzívne rozmnožovanie rastlín, najmä tých s pomalým prirodzeným rastom alebo ohrozených druhov, využíva technika rozmnožovania in vitro, teda v laboratórnych podmienkach z malých častí rastliny. Tento prístup je dôležitý pre masovú produkciu alebo záchranu ohrozených druhov.
- Explantát: Malá časť rastliny (list, stonka, bunka, protoplast) odobraná z materskej rastliny.
- Totipotencia: Vlastnosť všetkých buniek rastliny, ktoré obsahujú kompletnú genetickú informáciu a za určitých podmienok môžu z nich vzniknúť kompletné rastliny.
- Dediferenciácia: Proces, pri ktorom sa špecializovaná bunka vracia do stavu, keď je opäť schopná deliť sa. Pri dediferenciácii môže vzniknúť kalus (skupina nediferencovaných buniek) alebo meristematické rastové centrá.
- Regenerácia: Z dediferencovaných buniek možno regenerovať celú rastlinu buď formou organogenézy (vznik výhonku a následne koreňa) alebo somatickej embryogenézy (vznik bipolárnej štruktúry s apikálnymi meristémami koreňa a stonky).Príkladom je africká fialka, z ktorej jedného listu možno technikou in vitro získať až osemdesiat rastlín. Takto sa rozmnožujú napríklad aj ohrozené mäsožravé rastliny.
Porovnanie Pohlavného a Nepohlavného Rozmnožovania: Výhody a Nevýhody
Oba spôsoby rozmnožovania majú svoje výhody a nevýhody, ktoré ovplyvňujú ich rozšírenie a význam pre prežitie druhu.
Pohlavné Rozmnožovanie
- Výhody:
- Genetická variabilita potomstva: Umožňuje jedincom lepšie sa prispôsobiť meniacim sa podmienkam prostredia a obsadiť širšiu ekologickú niku.
- Odolnosť populácie: Znižuje pravdepodobnosť, že celá populácia bude citlivá voči jednému patogénu alebo náhlej zmene prostredia.
- Záloha genómu: Diploidný stav genómu poskytuje "zálohu" v prípade mutácie jedného génu, keďže je k dispozícii druhá funkčná kópia.
- Nevýhody:
- Pomalosť: Relatívna pomalosť v porovnaní s nepohlavným rozmnožovaním, pretože jedince sa môžu pohlavne rozmnožovať až po dosiahnutí pohlavnej dospelosti.
- Nižší počet jedincov: Často produkuje menší počet jedincov.
- Zložitosť: Vyžaduje zložitejší aparát a správanie (napr. opelenie), čo môže robiť jedince zraniteľnejšími.
- Zrieďovanie genetického materiálu: Dochádza k zrieďovaniu vlastného genetického materiálu.
Nepohlavné Rozmnožovanie
- Výhody:
- Rýchlosť: Rýchla kolonizácia stabilného prostredia.
- Energetická výhodnosť: Energeticky výhodnejšie, keďže nevyžaduje tvorbu pohlavných buniek a opelenie.
- Genetická zhodnosť: Genetická zhodnosť s rodičom, čo je výhodné v nemenných podmienkach, kde sú existujúce adaptácie už optimálne.
- Nevýhody:
- Nízka genetická variabilita: Znižuje schopnosť prispôsobenia sa zmenám prostredia.
- Zraniteľnosť populácie: Celá populácia je zraniteľná voči patogénom alebo nepriaznivým podmienkam, ktoré by mohli zničiť geneticky homogénne jedince.
Vyššie rastliny sa pohlavne rozmnožujú len za priaznivých vonkajších podmienok. Naopak, pri zhoršených podmienkach prechádzajú na pohlavný cyklus. Kým zo živočíchov sa vegetatívne rozmnožujú len vývojovo najnižšie druhy, z rastlín sa mnohými spôsobmi vegetatívne rozmnožujú aj vyššie rastliny.
Evolučný Význam a Diverzita Rastlín
Rastliny mali a majú z hľadiska evolúcie života na Zemi kľúčovú úlohu. Ich význam spočíva vo fotosyntéze, čo je dej, pri ktorom by ťažko mohol vzniknúť život v takej podobe, aký ho poznáme. Fotosyntéza je zdrojom takmer všetkých organických látok, ktoré vznikajú prirodzeným spôsobom, teda bez zásahu ľudskej technickej činnosti, a predstavuje autotrofný spôsob výživy všetkých zelených rastlín. Voda je súčasťou všetkých rastlinných buniek a je pre život rastliny nevyhnutná, dôležitým rozpúšťadlom a zdrojom vodíka a kyslíka pre fyziologické procesy. Základ bunkového dýchania rastlín je analogický so živočíchmi, no abiotický stres spôsobil vznik alternatívnych metabolických dráh. Špecifická oxidáza umožňuje termogenézu produkciou tepla na úkor syntézy ATP. Klíčiace semená zas dokážu vďaka glyoxylátovému cyklu konvertovať zásobné lipidy na sacharidy. Pohyby rastlín plnia inú funkciu ako pohyb živočíchov. Živočíchy si pohybom zabezpečujú potravu, obranu a reprodukciu. Pohyby rastlín majú rôznu formu a zabezpečujú rastline napríklad vhodné postavenie voči svetlu alebo uvoľňovanie semien.
Prispôsobenie sa Suchozemskému Prostrediu
Telo rastlín je tvorené z veľkého množstva buniek upravených pre určité životné funkcie. Nižšie rastliny majú telo tvorené z buniek, ktoré nie sú diferencované a vykonávajú všetky životné funkcie. S prechodom rastlín na suchú zem bol nevyhnutný vznik špecializovaných orgánov - koreň, zabezpečujúci príjem a rozvoz vody, stonka, zabezpečujúca mechanickú pevnosť a rozkonárovanie rastliny, list, primárne určený na fotosyntézu. Adaptácia na suché prostredie favorizovala rastliny s diferencovanou stavbou tela, čo viedlo k vzniku kormusu. Tento systém pravých orgánov a cievnych zväzkov zabezpečil rozvoz živín aj mimo vody.
Fylogenetické Línia a Ich Špecifiká
Nižšie rastliny sú fylogeneticky najstaršou vetvou eukaryotických rastlín. Z hľadiska rozmnožovania tvorí podríšu rastlín dvojica hlavných línií.
- Výtrusné rastliny: Predstavujú kľúčový fylogenetický medzistupeň pri osídľovaní pevniny. Svoj názov dostali podľa nepohlavného rozmnožovania pomocou spór. Adaptácia na suchozemské prostredie u nich viedla k vývoju krycích a vodivých pletív i diferenciácii pravých orgánov.
- Nahosemenné rastliny: Predstavujú dôležitý evolučný prechod ku krytosemenným druhom. Ich hlavným znakom je tvorba semien, ktoré ležia voľne odkryté na semenných šupinách, a keďže nemajú semenník, z botanického hľadiska netvoria skutočné plody. Vďaka vzniku peľovej trubice sa pri rozmnožovaní úplne oslobodili od vodného prostredia.
Krytosemenné Rastliny: Vrchol Evolúcie
Skupina pravých dvojklíčnolistových rastlín (eudikotov) predstavuje absolútny vrchol evolučného úspechu krytosemenných rastlín. Od starobylých línií ich spoľahlivo odlišuje inovatívny trikolpátny peľ vybavený tromi klíčiacimi otvormi. Pred vznikom pravých dvojklíčnolistových rastlín sa odštiepili bazálne vetvy a magnoliidy. Tieto prastaré evolučné línie si dodnes zachovali primitívne znaky, ako sú jednoduchšie cievice, špirálovité kvety a monokolpátny peľ s jedným otvorom.
Rozmanitosť Krytosemenných Rastlín: Dvojklíčnolistové a Jednoklíčnolistové
Fylogenetický strom vetvy pravých dvojklíčnolistových rastlín začína bazálnymi radmi s množstvom jedovatých druhov a plynule prechádza k jadrovým eudikotom.
- Superrosidy a rosidy: Predstavujú evolučný vrchol krytosemenných rastlín a zahŕňajú väčšinu našich poľnohospodárskych či lesných druhov. Ich fylogenetický strom začína bazálnymi radmi ovocných krov a sukulentov so špecifickou CAM fotosyntézou. Vývojová línia následne prechádza cez hospodársky dôležitý vinič k rozsiahlej COM vetve fabidov. Ricín obyčajný je mohutná tropická bylina s typickými dlaňovitými listami a semenami pripomínajúcimi fazuľu. Tieto semená ukrývajú mimoriadne toxický proteín ricín, ktorý na bunkovej úrovni nevratne blokuje syntézu bielkovín. Po technologickom spracovaní a denaturácii jedu sa však zo semien získava cenný ricínový olej. Fabidová vetva tvorí jednu z najdôležitejších fylogenetických línií na Zemi. Jej kľúčovou súčasťou je vetva rastlín schopných viazať atmosférický dusík vďaka symbióze s koreňovými baktériami. Do tejto gigantickej skupiny patria bôbotvaré byliny s typickým plodom strukom a vysokým obsahom bielkovín. Podľa modernej fylogenetiky APG IV ukrýva rad ružotvarých obrovské evolučné prekvapenie. Analýza DNA totiž potvrdila, že okrem klasických ruží a ovocných stromov s dužinatými plodmi sem patria aj zdanlivo úplne nesúvisiace druhy. Malvidová vetva je druhou hlavnou líniou rosidov, ktorá sa vyznačuje špecifickými chemickými a morfologickými adaptáciami. Nájdeme tu okrasné muškáty, javory, exotické citrusy, ale aj dreviny poskytujúce bavlnu či kakao. Mimoriadne dôležitou skupinou sú aj kapustovité rastliny.
- Superasteridy a asteridy: Bazálne línie superasteridov tvoria obrovskú skupinu pravých dvojklíčnolistových rastlín, pre ktorú je typická produkcia špecifických iridoidov. Počas evolúcie si vybudovali extrémne stratégie na prežitie v nehostinných podmienkach. Do tejto vetvy patria poloparazitické dreviny, suchomilné kaktusy, čajovníky aj dôležité poľnohospodárske plodiny. Lamiidová vetva je kľúčovou líniou asteridov, typickou prítomnosťou jediného vajíčkového obalu a tvorbou iridoidov. Z biochemického hľadiska vyniká masívnou produkciou alkaloidov a prchavých silíc. Do tejto skupiny radíme hospodársky dôležité ľuľkovité plodiny, kávovníky či obľúbené hluchavkovité byliny. Campanulidová vetva predstavuje spolu s lamiidmi evolučný vrchol asteridov. Tieto rastliny charakterizuje jediný vajíčkový obal a prítomnosť špecifických iridoidov. Do tejto fylogenetickej línie patrí mimoriadne úspešná čeľaď astrovitých so súkvetím úbor, ale aj mrkvovité rastliny tvoriace okolíky.
- Jednoklíčnolistové rastliny: Skupina jednoklíčnolistových rastlín predstavuje významnú evolučnú líniu, odlišujúcu sa od dvojklíčnolistových zásadnými anatomickými znakmi. Typický je pre ne jediný klíčny list, vedľajšia koreňová sústava (homorízia) a stonka bez druhotného hrubnutia. Fylogenetický strom tejto skupiny zahŕňa jedovaté jesienky, cibuľoviny i vzácne orchidey. Rad lipnicotvaré predstavuje kľúčovú skupinu jednoklíčnolistových rastlín, do ktorej patrí hospodársky najvýznamnejšia čeľaď tráv. Tieto vetrom opelivé druhy sú typické dutou stonkou (steblom) a suchými plodmi, ktoré v poľnohospodárstve poznáme ako obilné zrná.
tags: #oplodnenie #a #vyvin #novej #rastliny