
Reprodukcia, čiže rozmnožovanie každého jedinca a jeho rast, sú neoddeliteľne spojené s delením buniek. Reprodukcia buniek je jednou zo všeobecných vlastností buniek, ktorá je pre život nevyhnutná. Materské bunky sa delia, reprodukujú, čím dávajú vznik novým dcérskym bunkám, ktoré sú geneticky zhodné s materskou bunkou. Tento fundamentálny proces je základom individuálneho vývinu, takzvanej ontogenézy mnohobunkového organizmu, a zároveň dáva vznik aj špecializovaným pohlavným bunkám. Výsledkom reprodukcie buniek sú nové generácie buniek a v širšom kontexte aj nových mnohobunkových organizmov. Reprodukcia je teda kľúčovou časťou života bunky, v ktorom sa bunky delia, rastú a vyvíjajú sa podľa genetickej informácie zapísanej v DNA, ktorú získali z materskej bunky. Okrem zabezpečenia rozmnožovania a rastu má bunkové delenie aj inú vitálnu úlohu, konkrétne zabezpečuje regeneráciu poškodených tkanív, orgánov a náhradu opotrebovaných buniek, čím prispieva k udržaniu celistvosti a funkčnosti organizmu. Z hľadiska vývoja je delenie buniek základom kontinuity, udržania nepretržitosti života a druhu, potvrdzujúc tak cyklickú povahu existencie. V súčasnej etape fylogenetického vývoja vznikajú nové bunky reprodukciou buniek už existujúcich, čiže ich delením.
Bunkový Cyklus: Rytmus Bunkového Života
Život bunky, ktorý predstavuje striedanie obdobia rastu a delenia, sa nazýva bunkový cyklus. Tento cyklus je cyklicky sa opakujúce delenie buniek, ktoré sa vo vhodných podmienkach neustále opakuje. Keď sa bunky po skončení bunkového delenia osamostatnia, okamžite začínajú prípravné fázy na ďalšie delenie buniek. Tieto prípravné procesy prebiehajú v interfáze bunkového cyklu, čo je obdobie, keď sa bunka nedelí. Súčasťou bunkového cyklu sú všetky procesy, ktoré sa dejú v bunke od okamihu vzniku dcérskej bunky po mitóze, až po jej opätovné rozdelenie. Rozdelenie cyklu na štyri fázy, ktoré v roku 1953 navrhli rádiobiológovia Alma Howard a Stephen Pelc, je dnes všeobecne prijaté a predstavuje štandardné poňatie priebehu bunkového cyklu. Hlavným kritériom tohto delenia cyklu je replikácia jadrovej DNA a vlastné rozdelenie bunky, teda mitóza.
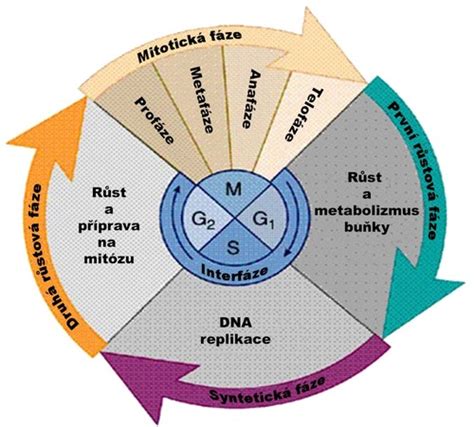
Bunkový cyklus eukaryotických buniek sa skladá z niekoľkých fáz:
Interfáza: Príprava na Delenie
Interfáza je najdlhšia fáza bunkového cyklu a je kľúčová pre prípravu bunky na delenie. Bunka v nej intenzívne rastie, plní svoje bežné metabolické funkcie a fyziologicky sa chystá na delenie. Začína sa bezprostredne po vzniku novej bunky a je ďalej rozdelená na tri podfázy:
G1-fáza (Postmitotická fáza): Táto fáza začína najčastejšie v okamihu rozdelenia po vzniku novej bunky. Bunka sa tu zotavuje z predošlého delenia a intenzívne rastie. Prebiehajú tu najmä rastové procesy, spojené so syntézou bielkovín, a zároveň sa syntetizuje RNA a dopĺňajú sa bunkové organely. Je to obdobie tvorby zásoby nukleotidov a enzýmov nevyhnutných pre neskoršie zdvojenie DNA. V G1-fáze sa nachádza aj hlavný kontrolný uzol bunkového cyklu, ktorý má kľúčovú regulačnú funkciu. V ňom sa bunkový cyklus zastavuje pri regulácii bunkového delenia v prípade nepriaznivých podmienok vonkajšieho prostredia, napríklad pri nedostatku živín, pri nevhodnej teplote, alebo pri nahromadení škodlivých látok. Toto zastavenie blokuje nielen priebeh G1-fázy, ale aj celého bunkového cyklu. Bunky, ktoré sa v mnohobunkovom organizme nedelia, napríklad neuróny alebo svalové bunky, sa nachádzajú v G1-fáze v tzv. G0-fáze. Toto pokojové štádium, nazývané kviescencia, je reverzibilné a bunka môže po čase opäť vstúpiť do aktívneho cyklu. Bunka v tomto štádiu žije, plní svoje špecifické funkcie v organizme a morfologicky sa diferencuje, ale nedelí sa. V mnohobunkových organizmoch sa v tejto fáze nachádza drvivá väčšina buniek. Ak bunka úspešne prejde kontrolným uzlom, vstupuje do energeticky mimoriadne náročnej S-fázy.
S-fáza (Syntetická fáza): V tejto fáze prebieha replikácia (syntéza DNA), ktorej výsledkom je presné zdvojnásobenie genetického materiálu. Počas syntetickej interfázy, ktorá je predmitotickou fázou, sa znásobujú genetické materiály v bunke. To znamená, že jednochromatidový chromozóm sa mení na dvojchromatidový, spojený v mieste prvotného delenia, čiže centroméry. Rovnako sa zdvojuje, replikuje, aj v ňom uložená DNA na dvojnásobok. Každý chromozóm je na konci tejto fázy tvorený dvomi identickými sesterskými chromatídami, ktoré sú fyzicky spojené v mieste centroméry. Bunka je v tomto momente fyziologicky tetraploidná ((2 \times 2n = 4n)).
G2-fáza (Premitotická fáza): Toto je záverečná časť interfázy, v ktorej sa bunka finálne chystá na delenie. Podobne ako v G1-fáze, aj tu pokračujú intenzívne syntetické procesy - najvýraznejšie sa syntetizuje RNA a bielkoviny, ako napríklad tubulín, nevyhnutný pre stavbu deliaceho vretienka. Zároveň prebieha delenie semiautonómnych organel, ako sú mitochondrie a plastidy, čomu predchádza replikácia ich vlastnej mimojadrovej DNA. Bunka ďalej rastie a pripravuje sa na delenie jadra a samotnej bunky.
M-fáza (Mitotická fáza): Vlastné Delenie Bunky
Táto fáza predstavuje samotné delenie bunky. Z časového hľadiska predstavuje samotné delenie bunky zhruba 5 - 10 % celkového trvania cyklu. Táto fáza sa skladá z dvoch logicky a fyziologicky odlišných, ale úzko nadväzujúcich procesov: delenia jadra - karyokinézy a delenia samotnej bunky - cytokinézy. V bežnej reči sa pojmom mitóza často zjednodušene označuje celé delenie bunky, avšak z hľadiska presnej terminológie mitóza pomenúva výlučne delenie jadra a genetického materiálu. Často sa stáva, že jadro sa viacnásobne rozdelí, ale bunka sa rozdelí iba raz, čoho výsledkom sú viacjadrové bunky - typické pre nižšie mikroskopické huby alebo pre priečne pruhované svalové vlákna.
Generačná Doba Bunky
Časové trvanie bunkového cyklu sa nazýva generačná doba bunky. Je daná geneticky a pre rozličné bunky je rozdielna - napríklad baktérie majú generačnú dobu asi 30 minút, zatiaľ čo prvoky niekoľko hodín. Hlavným faktorom dĺžky života bunky sú vhodné životné podmienky, ako je teplota, dostatok živín a iné. Pokiaľ sa zastaví prísun živín, hlavný kontrolný uzol zablokuje delenie bunky a trvanie bunkového cyklu sa predĺži po určitú kritickú hranicu. Ak sa prísun živín neobnoví, bunka hynie.
Chromozómy: Nositelia Genetickej Informácie
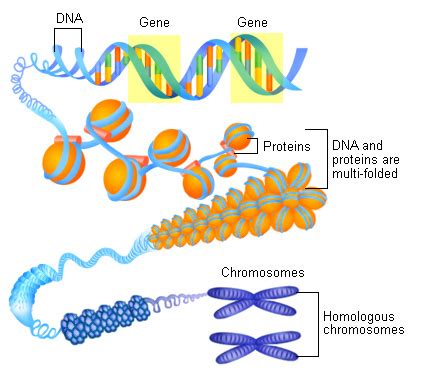
Chromozóm sa považuje za základnú jednotku bunkového delenia a je kľúčovým prvkom pre prenos genetickej informácie. Sú tvorené nukleoproteínovými vláknami a ich počet, tvar a veľkosť sú pre každý druh organizmu charakteristické a relatívne stále. Pozdĺžne rozdelený chromozóm, ako ho vidíme počas delenia, pozostáva z dvoch chromatíd, ktoré sú viditeľné ako dvojice paralelných chromonémov. Tieto chromatidy sú spojené v mieste prvotného delenia, takzvanej centroméry. Centroméra je miesto prvotného zúženia chromozómu, nazývaného aj primárna konstrikcia, a tento úsek chromozómu má dôležitú úlohu pri pohybe chromozómov počas delenia bunky. Na chromozómoch sa rozlišujú dve ramená, ktoré sú spojené práve v mieste centroméry. Niektoré chromozómy majú aj druhotné zúženie, takzvanú sekundárnu konstrikciu, kde sa nachádza prívesok - satelit, spravidla oddelený organizátorom jadierka.
Kompletný súbor chromozómov v bunke sa nazýva sada. U človeka má sada 23 chromozómov. V telových bunkách, ktoré vznikli delením zygoty (zygota vznikla splynutím dvoch pohlavných buniek - samčej a samičej), sa nachádzajú dve sady chromozómov. Telové, čiže somatické bunky sú diploidné, označujeme ich 2n, a majú dvojnásobný počet chromozómov. U človeka telové bunky obsahujú 46 chromozómov. V somatických bunkách sa chromozómy vyskytujú v pároch, sú to párové homologické chromozómy, z ktorých jeden pochádza od otca a druhý od matky. Homologické chromozómy nesú informácie o tých istých znakoch, sú to dve kópie tých istých chromozómov. U mužov sú výnimkou iba pohlavné chromozómy XY, ktoré sú heterologické. Naopak, pohlavné bunky majú polovičný počet chromozómov, sú haploidné a označujeme ich n.
Bunkové Delenie: Typy a Mechanizmy
Sú známe dva základné spôsoby delenia buniek: mitóza - nepriame delenie a meióza - redukčné delenie. Existuje aj zriedkavý spôsob delenia buniek - amitóza, čo je priame delenie. Každý z týchto spôsobov slúži inému účelu v živote organizmu.
Mitóza: Nepriame Delenie
Mitóza je najčastejší spôsob delenia eukaryotických buniek. Materská bunka sa mitózou rozdelí na dve rovnocenné dcérske bunky, ktoré sa geneticky zhodujú s materskou bunkou. Táto genetická zhoda je kľúčová pre rast, vývoj a regeneráciu tkanív. Mitóza zabezpečuje presné rozdelenie genetického materiálu nachádzajúceho sa v chromozómoch do dcérskych buniek, čím sa zachová genetická zhoda materských a dcérskych buniek. Presné rozdelenie chromozómov do dcérskych buniek prebieha za pomoci mitotického aparátu, ktorý tvoria dve súčasti - centriol a deliace vretienko. Centriol je stála štruktúra bunky uložená na povrchu jadrového obalu. Deliace vretienko je sústava mikrotubulov, ktorá vznikla už na začiatku mitózy a po jej skončení zaniká; nie je to teda stála štruktúra. Vlastnému deleniu bunky predchádzajú intenzívne syntetické procesy v interfáze bunkového cyklu, najmä zdvojenie DNA a celej hmoty chromozómu v S-fáze.
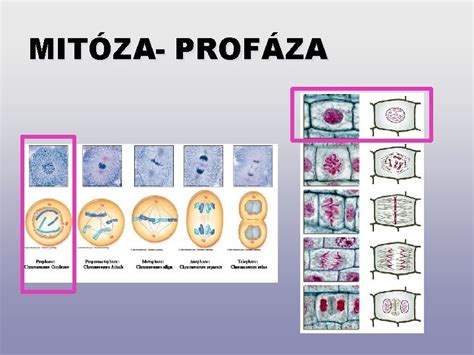
Mitóza prebieha v nasledujúcich fázach:
Profáza: V tejto úvodnej fáze sa chromozómy skracujú a hrubnú - špiralizujú sa, a tak sa stávajú rozlíšiteľné a viditeľné pod mikroskopom. Sú na nich rozlíšené dve chromatídy, ktoré ale zostávajú spojené v mieste centroméry. Jadrová membrána sa rozpúšťa a zaniká, jadierko tiež mizne a objavuje sa deliace vretienko. Centriol sa rozdelí na dve časti, ktoré sa presúvajú k protiľahlým pólom bunky, čím sa vytvárajú centrá deliaceho vretienka.
Metafáza: V metafáze vrcholí špiralizácia, chromozómy sa maximálne skracujú, a preto sú najlepšie pozorovateľné, dajú sa počítať a identifikovať. Ich fotografovaním sa robí takzvaný karyotyp - presný obraz chromozómov bunkového jadra. Dvojchromatidové chromozómy sa v tejto fáze zaraďujú do centrálnej - rovníkovej (ekvatoriálnej) roviny bunky, kde utvoria hviezdicovitý útvar, nazývaný monaster. Deliace vretienko sa mikrotubulami pripája na jednom konci k centriolám a na druhom k centroméram chromozómov. Dvojchromatidový chromozóm sa postupne pripravuje na rozdelenie na dve dcérske chromatidy.
Anafáza: Po úplnom rozdelení centroméry na dve časti dochádza k skracovaniu mikrotubúl deliaceho vretienka. Týmto mechanizmom sa dcérske chromatidy každého chromozómu oddelia a rozchádzajú na protiľahlé póly bunky. Chromozómy sa priťahujú k centriolám vďaka skracovaniu mikrotubulov deliaceho vretienka. Pri centriolách sa na každom póle sústreďuje znova diploidný počet chromozómov, pričom každý chromozóm je už jednochromatidový.
Telofáza: V telofáze deliace vretienko zaniká. Jednochromatidové dcérske chromozómy sa dešpiralizujú, to znamená, že sa menia na dlhé vlákna, a okolo nich sa vytvorí nová jadrová membrána. Objavuje sa jadierko, ktoré bolo v profáze zaniknuté. Obnovením štruktúry jadra končí karyokinéza, delenie jadra, a nasleduje cytokinéza - rozdelenie materskej bunky na dve samostatné dcérske bunky. Tieto dcérske bunky majú ten istý počet chromozómov ako mala materská bunka. Znamená to, že ak bola materská bunka diploidná (2n), budú diploidné (2n) aj dcérske bunky. Podstata mitózy eukaryotickej bunky spočíva v tom, že bunka sa rozdelí raz a raz sa aj replikuje (zdvojí) DNA.
Cytokinéza: Delenie Cytoplazmy
Cytokinéza predstavuje samotné fyzické rozdelenie cytoplazmy a bunkových organel, čím z jednej materskej bunky definitívne vzniknú dve samostatné dcérske bunky. Mechanizmus tohto delenia priamo závisí od prítomnosti bunkovej steny:
U živočíšnych buniek: Delenie prebieha zaškrtením cytoplazmatickej membrány od vonkajšieho povrchu do stredu (dostredivo, centripetálne). Tento proces zabezpečuje kontraktilný prstenec tvorený z bielkovín aktínu a myozínu, ktorý sa postupne sťahuje.
U rastlinných buniek: Pre prítomnosť pevnej bunkovej steny sa bunka nemôže jednoducho zaškrtiť. Namiesto toho sa v strede bunky začnú hromadiť vezikuly (mechúriky) z Golgiho aparátu. Tieto vezikuly sa spájajú a vytvárajú priehradku - plazmatickú platničku, ktorá postupne rastie od stredu k okrajom bunky, až kým ju nerozdelí na dve samostatné bunky. Vznik plazmatickej platničky možno pozorovať iba elektrónovým mikroskopom. Po vytvorení nových jadier v telofáze sa rastlinné bunky v procese cytokinézy vytvárajú z vezikúl, ktoré sú oddelené od diktyozómov v centrálnej rovine.
Mitóza: Úžasný bunkový proces, ktorý využíva delenie na množenie! (Aktualizované)
Amitóza: Priame Delenie
Amitóza je pomerne zriedkavý spôsob bunkového delenia. Takto sa delia zrejme len patogénne bunky a bunky vyžadujúce urýchlenú reparáciu opotrebovaných alebo poškodených častí orgánov. V priebehu amitózy sa chromozómy nezdvojujú a nevytvára sa deliaci aparát. Jadro sa postupne zaškrcuje na dve časti, pričom zákonite nie je zaručené rovnomerné rozdelenie genetického materiálu. Po rozdelení jadra sa rozdelí aj cytoplazma a vznikajú dve dcérske bunky, ktoré však nemusia byť geneticky rovnocenné.
Meióza: Redukčné Delenie
Meióza je osobitný spôsob delenia buniek, pri ktorom nastáva redukcia - zmenšenie počtu chromozómov na polovicu. Preto je meióza jediný možný spôsob vzniku pohlavných buniek, takzvaných gamét. U živočíchov sú to vajíčka a spermie, u vyšších rastlín spermatické bunky v peľovom zrnku a vajcové bunky. Pohlavné bunky musia mať polovičný počet chromozómov v porovnaní s telovými bunkami, pretože ak by mali rovnaký počet chromozómov ako telové bunky, t.j. diploidný (2n), ich splynutím by vznikol jedinec so 4n počtom chromozómov. Počet chromozómov by z generácie na generáciu rástol, čo nie je biologicky udržateľné. Meióza prebieha v dvoch po sebe nasledujúcich deleniach: prvom meiotickom delení, ktoré je heterotypické, a druhom meiotickom delení, ktoré je homeotypické. Meióza prechádza podobne ako mitóza štádiom profázy, metafázy, anafázy a telofázy, ktoré odrážajú zmeny v jadre a na chromozómoch, avšak s dôležitými špecifikami.
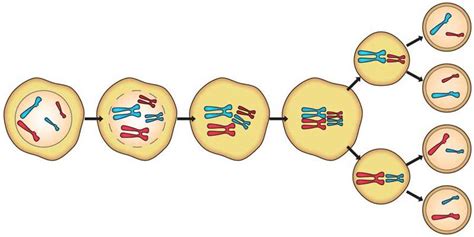
Prvé Meiotické Delenie (Heterotypické Delenie)
Prvé meiotické delenie je charakteristické redukciou počtu chromozómov na polovicu, preto sa nazýva heterotypické (hetero = iný).
Profáza I: Priebeh profázy heterotypického delenia je na rozdiel od mitózy zložitejší a podstatne dlhší. Začína sa po skončení interfázy, ktorej výsledkom je zdvojenie - replikácia DNA a vytvorenie dvojchromatidových chromozómov. V profáze I sa chromozómy stávajú viditeľné a homologické chromozómy sa párujú, vytvárajú dvojice, takzvané bivalenty. Keďže každý z homologických chromozómov je zdvojený, dvojchromatidový, v každom bivalente možno rozlíšiť štyri chromatidy homologických chromozómov. Vznik bivalentov je podmienený tvorbou špecifickej bielkoviny. Susediace chromatidy homologických chromozómov sa spravidla prekrížia, čo sa nazýva crossing-over, a vymenia si navzájom genetický materiál. Takto vznikajú chromozómy z časti otcovského aj materského pôvodu, čo prispieva k genetickej variabilite. Na konci profázy I sa homologické chromozómy oddeľujú, jadrová membrána zaniká a vytvára sa deliace vretienko.
Metafáza I: V tejto fáze sa bivalenty, tvorené pármi homologických dvojchromatidových chromozómov, radia do ekvatoriálnej roviny bunky. Deliace vretienko sa pripája k centroméram každého homologického chromozómu v páre, pričom jeden chromozóm z páru smeruje k jednému pólu a druhý k opačnému.
Anafáza I: V anafáze I sa homologické chromozómy, ktoré sú stále zdvojené, rozchádzajú na protiľahlé póly bunky. Týmto procesom nastáva redukcia počtu chromozómov na polovicu. Podstatný rozdiel oproti anafáze mitózy je v tom, že v anafáze meiotického delenia putujú k pólom celé dvojchromatidové, teda zdvojené chromozómy, nie jednotlivé chromatidy.
Telofáza I: V telofáze I sa chromozómy dešpiralizujú a vytvorí sa jadrová membrána okolo dvoch vznikajúcich jadier. Vznikajú dve dcérske jadrá - to je karyokinéza, a následne nastáva cytokinéza, ktorej výsledkom sú dve dcérske bunky s polovičným počtom chromozómov, pričom každý chromozóm je stále dvojchromatidový.
Po prvom meiotickom delení nastáva krátka interfáza, v ktorej však nedochádza k ďalšej replikácii DNA. Po tejto krátkej interfáze nasleduje druhé meiotické delenie.
Mitóza: Úžasný bunkový proces, ktorý využíva delenie na množenie! (Aktualizované)
Druhé Meiotické Delenie (Homeotypické Delenie)
Druhé meiotické delenie sa nazýva homeotypické (homo = rovnaký), pretože počet chromozómov sa v jeho priebehu nemení a je v princípe podobné ako normálna mitóza, avšak prebieha už v bunkách s haploidným počtom chromozómov. Pokračuje v obidvoch bunkách, ktoré vznikli po prvom meiotickom delení. Má štyri fázy podobne ako mitóza, ale keďže má svoje špecifiká, tieto fázy sa označujú ako Profáza II., Metafáza II., Anafáza II. a Telofáza II.
Profáza II: Jadrová membrána sa opäť stráca, chromozómy sa zviditeľňujú a vytvára sa nové deliace vretienko v každej z dvoch dcérskych buniek.
Metafáza II: Dvojchromatidové chromozómy sa radia do ekvatoriálnej roviny v oboch bunkách. Deliace vretienko sa pripája k centroméram chromozómov.
Anafáza II: V tejto fáze sa dvojchromatidové chromozómy pozdĺžne rozdelia na dve chromatidy. Chromatidy sa oddeľujú a putujú k protiľahlým pólom bunky, stávajúc sa jednochromatidovými dcérskymi chromozómami. Podstatný rozdiel oproti mitóze spočíva v tom, že v anafáze prvého meiotického delenia sa k pólom presúvajú celé dvojchromatidové chromozómy, zatiaľ čo v anafáze II meiózy sa oddeľujú a presúvajú jednotlivé chromatidy.
Telofáza II: Okolo zoskupených jednochromatidových dcérskych chromozómov na póloch buniek sa vytvára nová jadrová membrána. Chromozómy sa dešpiralizujú a jadierka sa objavujú. Nasleduje cytokinéza v oboch bunkách. Konečným výsledkom meiózy sú štyri bunky - gaméty s haploidným počtom jednochromatidových chromozómov (n). Z jednej diploidnej (2n) bunky sa teda vytvárajú štyri haploidné (n) bunky. Je nutné, aby gaméty mali polovičný (n) počet chromozómov, pretože pri ich splývaní (oplodnení) vzniká zygota, ktorá bude mať opäť diploidný (2n) počet chromozómov. Zygota sa ďalej mitotický delí, čím sa zabezpečí, že všetky telové bunky nového organizmu budú mať diploidný (2n) počet chromozómov, čím sa udržuje stabilný počet chromozómov v druhu.
Regulácia Bunkového Cyklu a Osud Buniek
Reguláciu bunkového cyklu zabezpečujú komplexné regulačné mechanizmy. Tieto mechanizmy riadia priebeh bunkového cyklu a na úrovni mnohobunkového organizmu zabezpečujú zodpovedajúci počet buniek vo všetkých tkanivách a orgánoch, čím udržiavajú homeostázu a správne fungovanie organizmu. Regulácia bunkového delenia je preto dôležitá pri bunkovej reprodukcii.
Najčastejšie je látkový, čiže chemický spôsob regulácie bunkového cyklu. Znamená to, že niektoré chemické látky majú stimulačný účinok, t.j. iniciujú bunkové delenie, a iné, naopak, spomaľujú, až zastavujú bunkové delenie, t.j. pôsobia inhibične. Všetky regulátory bunkového delenia ovplyvňujú priebeh bunkového cyklu predovšetkým v G1-fáze, kde je hlavný kontrolný uzol bunkového cyklu. Medzi látky, ktoré spomaľujú až zastavujú bunkové delenie, patria cytostatiká, ktoré sa úspešne využívajú pri liečbe nádorových ochorení, pretože bránia nekontrolovanému deleniu rakovinových buniek.
Niektoré bunky rastlinných pletív, napríklad sitkovice, a živočíšnych tkanív, ako sú bunky mozgu (neuróny) alebo erytrocyty, majú geneticky trvalo zablokovanú schopnosť deliť sa. Znamená to teda, že u týchto buniek sa bunkový cyklus preruší. Bunky, ktoré natrvalo stratili schopnosť delenia, majú kontrolný uzol v G1-fáze natrvalo zablokovaný. V mnohobunkových organizmoch teda bunkový cyklus riadia regulačné mechanizmy, ktoré zabezpečujú zodpovedajúci počet buniek vo všetkých tkanivách a orgánoch. Regulácia bunkového cyklu je jedným z hlavných mechanizmov zabezpečujúcich celistvosť mnohobunkového organizmu.
Reguláciu bunkového cyklu môžu výrazne ovplyvniť aj vírusy. Tieto, pri spolupôsobení vnútorných, ako aj vonkajších rizikových faktorov, môžu za istých okolností spôsobiť nekoordinované delenie buniek, a tak vyvolávať tvorbu nádorov, čo podčiarkuje zložitosť a zraniteľnosť regulačných procesov.

Život bunky však nie je obmedzený výlučne na rast a reprodukciu prostredníctvom bunkového cyklu. Podľa prítomnosti extracelulárnych faktorov a fyziologického stavu prechádza bunka aj rôznymi inými procesmi:
- Kviescencia (G0-fáza): Je to pokojové štádium, ktoré je reverzibilné. Bunka môže po čase opäť vstúpiť do aktívneho cyklu. V tomto štádiu bunka žije, plní svoje špecifické funkcie v organizme a morfologicky sa diferencuje, ale nedelí sa. V mnohobunkových organizmoch sa v tejto fáze nachádza drvivá väčšina buniek. Hoci má každá bunka schopnosť deliť sa, nie všetky bunky mnohobunkového organizmu sa delia. Pri rastlinách sú to bunky nedelivých (trvácich) pletív, pri živočíchoch sú to v dospelosti bunky takmer všetkých tkanív.
- Diferenciácia: Tento proces vedie k vzniku funkčne špecializovaných a tvarovo odlišných buniek, ktoré tvoria tkanivá a pletivá. Pri delení dochádza k funkčnému a štruktúrnemu rozmnožovaniu, čo je bunková diferencia.
- Senescencia: Je to stav, kedy sa bunky už z G0-fázy nemôžu vrátiť do bunkového cyklu. Často sa označuje ako terminálna diferenciácia, čo znamená, že bunka dosiahla svoj konečný, nedelivý stav a vykonáva špecifické funkcie až do svojho zániku.
- Apoptóza: Predstavuje programovanú bunkovú smrť. Je to regulovaný proces, ktorý je nevyhnutný pre správny vývoj organizmu, elimináciu poškodených alebo nepotrebných buniek a udržiavanie tkanivovej homeostázy.
Všetky ostatné životné funkcie vykonáva bunka bez prerušenia, aj keď sa nenachádza v aktívnom bunkovom cykle, čo ukazuje na dynamickú a komplexnú povahu bunkového života.
tags: #delenie #reprodukcia #buniek