
Úvod: Fenomén dvojitého oplodnenia
Rozmnožovanie je základným kameňom existencie všetkých živých organizmov, zabezpečujúcim kontinuitu druhu a hnacím motorom evolúcie. Tento komplexný proces, zahŕňajúci pestrú škálu etologických, fyziologických a morfogenetických dejov, sa uplatňuje v rozličných formách v závislosti od organizmu. V rastlinnej ríši dosahuje proces sexuálneho rozmnožovania vrchol v podobe fascinujúceho javu známeho ako dvojité oplodnenie, ktoré je výsadou krytosemenných rastlín a kľúčom k ich dominancii na našej planéte. Krytosemenné rastliny (Angiospermae, Angiospermatophyta) sú rozsiahly taxón rastlín, charakteristický tým, že ich semená sú ukryté v plode. Vzniku plodu predchádza obojpohlavný alebo jednopohlavný kvet, čo je zásadný rozdiel oproti nahosemenným rastlinám, ktorým typické kvety s obalmi a bliznou chýbajú. Tento článok sa ponorí do podrobností vývoja pohlavných orgánov a samotného procesu dvojitého oplodnenia u krytosemenných rastlín, objasní jeho evolučný význam a poskytne porovnanie s rozmnožovaním nahosemenných rastlín.
Vývin samčieho gametofytu: Peľové zrno a jeho cesta k oplodneniu
Samčie reprodukčné orgány rastlín sú tyčinky. V tyčinkách sa nachádzajú peľové komôrky, v ktorých sa tvoria peľové zrná. Dozrievanie peľového zrna, teda vznik samčieho gametofytu, je zložitý proces. Peľové zrná vznikajú v peľnici po redukčnom delení. Vnútri juvenilného mikrosporangia sa nachádza peľotvorné pletivo - archespór. Z neho sa diferencuje výstelkové pletivo - tapetum a sporogénne pletivo. Redukčným delením buniek sporogénneho pletiva vzniknú tetrády haploidných mikrospór.
Bunková stena vyvíjajúcich sa peľových zŕn je v tomto období tvorená exinou a vnútornou intinou. U borovice a ďalších druhov ihličnatých rastlín exina odstáva a vytvárajú sa vzdušné vaky, ktoré umožňujú prenos peľových zŕn na veľké vzdialenosti. Hoci krytosemenné rastliny takýmito výraznými vzdušnými vakmi nedisponujú v rovnakej miere, ich peľové zrná sú prispôsobené na rôznorodé formy prenosu.
Braun a kol. (1976) rozlišujú primárne, sekundárne a terciálne delenie v rámci formovania sa samčieho gametofytu. Mikrospóry sa podobne ako megaspóry začínajú deliť ešte v sporangiu. Vzniknuté bunky majú haploidné jadro, ktoré sa mitoticky rozdelí a vzniknú dve haploidné jadrá: vegetatívne (vyživovacie) a generatívne (rozmnožovacie). Neskôr sa generatívne jadro ešte raz rozdelí a vznikajú dve tzv. spermácie, ktoré sú tiež označované ako samčie neobrvené bunky. Práve tieto spermatické jadrá sa zúčastnia oplodnenia ako samčie pohlavné bunky (gaméty). V čase zrelosti sa peľové zrnká krytosemenných rastlín skladajú z jednej vegetatívnej bunky, ktorá zabezpečuje rast peľovej trubice, a dvoch samčích generatívnych buniek.

Vývin samičieho gametofytu: Zárodočný miešok a vajíčko
Samičie pohlavné orgány krytosemenných rastlín, piestik, obsahujú vajíčka. Samičí gametofyt sa vyvíja vo vajíčku semenníka. Vajíčka u semenných rastlín vznikajú na plodolistoch, ktoré sa u nahosemenných a krytosemenných rastlinách odlišujú. Plodolisty krytosemenných rastlín zrastajú a tvoria piestik (gynaeceum). Na každom plodoliste sa nachádza zárodočné pletivo (placenta), kde sa vytvárajú vajíčka.
Vajíčko u rastlín je mnohobunkový útvar, ktorý pokrývajú obaly - integumenty. Obaly vajíčka - integumenty - sú dva, vzácnejšie jeden, a obrastajú celé vajíčko. Na vrchole však zostáva nezrastený otvor - peľový vchod (mikropyla), ktorý umožňuje samčím gamétam preniknúť do vajíčka a oplodniť vajcovú bunku. Vajíčko je chránené plodolistami, ktoré niekedy zrastajú a vytvárajú piestik. V jednom piestiku sa môže nachádzať aj viac vajíčok. Vo vnútri vajíčka sa nachádza pletivo nucelus, ktorého jedna bunka - materská bunka zárodočného mieška - sa zväčšuje a redukčne sa delí. Po tomto meiotickom delení vzniknú štyri haploidné bunky. U väčšiny rastlín tri z týchto buniek zanikajú a zostane len jedna bunka, ktorá sa nazýva jednobunkový mladý zárodočný miešok. U niektorých rastlín však môžu zostať dve alebo výnimočne všetky štyri haploidné bunky (bispórický, resp. tetraspórický typ).
V tejto zostávajúcej spóre prebehnú tri mitotické delenia a vznikne tak 8-jadrový, 7-bunkový útvar, označovaný ako zrelý zárodočný miešok krytosemenných rastlín. V jeho strede vzniká diploidné centrálne jadro (často tvorené splynutím dvoch polárnych jadier). K hornému koncu útvaru, v blízkosti mikropyly, sa premiestni vajcová bunka (oosféra) sprevádzaná dvoma synergidami. Na opačnom, chalazálnom konci zárodočného mieška sa nachádzajú tri antipódy. Samičí gametofyt krytosemenných rastlín je teda zredukovaný len na sedem buniek (zárodočný vak). Oosféra je samičia gaméta, pripravená na oplodnenie.
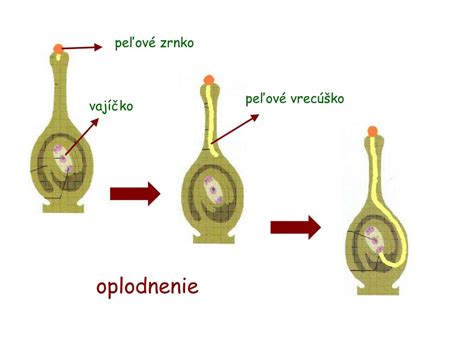
Opelenie u krytosemenných rastlín: Most k splynutiu
Opelenie (pollinatio) je prenesenie peľu z tyčinky na bliznu piestika. Peľové zrnko sa musí nejakým spôsobom dostať na bliznu piestika - za pomoci vetra (vetroopelivé rastliny), prostredníctvom živočíchov, najčastejšie hmyzu (včely), ale aj vtákov (kolibríky) alebo malých cicavcov. U niektorých rastlín môže peľ preniesť aj voda. Podľa pôvodu peľu rozlišujeme samoopelenie (autogamiu), ak je peľ prenesený z tyčinky na piestik toho istého kvetu, a cudzoopelenie (allogamia), ak je peľ prenesený na cudzí kvet.
Rastliny sa samoopeleniu bránia rôznymi mechanizmami, ako je autoinkompatibilita (biochemický blok), časový nesúlad dozrievania orgánov (napríklad proterandria, keď dozrievajú najprv tyčinky, alebo proterogynia, keď dozrieva najprv blizna) alebo priestorové usporiadanie kvetov. Dômyselný priestorový mechanizmus na podporu cudzoopelenia využíva napríklad prvosienka jarná. Jednotlivé rastliny tvoria buď kvety s dlhými čnelkami a krátkymi tyčinkami, alebo presne naopak, čo priamo núti hmyz prenášať peľ výhradne medzi rôznymi jedincami. Špecifickým prípadom samoopelenia je kleistogamia (skrytoopelivosť), pri ktorej dochádza k opeleniu ešte v púčiku, teda ešte predtým, ako kvet rozkvitne.
Ak na bliznu dopadne správny peľ, vlhká blizna, prítomný vápnik, bór a fytohormóny ho okamžite stimulujú ku klíčeniu. Peľové zrnko začne klíčiť a vytvorí sa peľová trubica (peľové vrecúško). Táto trubica prerastá cez čnelku až do semenníka. Do peľovej trubice sa presúvajú vegetatívne jadro a obe už vytvorené spermatické jadrá (spermácie). Peľová trubica prerastá až k obalom vajíčka, typicky cez mikropylárny otvor, čím umožní samčím gamétam preniknúť do vajíčka.
Proces opeľovania a oplodnenia 🌷| Postup krok za krokom
Kľúčový moment: Dvojité oplodnenie
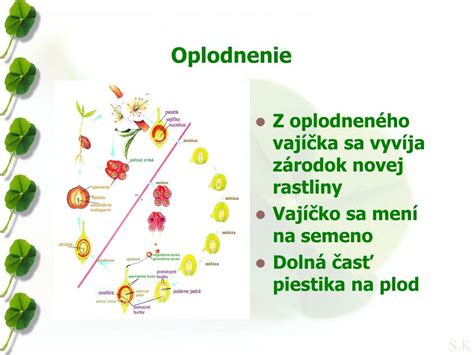
Dvojité oplodnenie je proces, ktorý sa vyskytuje výhradne u krytosemenných rastlín a predstavuje významný evolučný krok. Tento proces, objavený ruským botanikom S. Navashinom, zahŕňa dve splynutia gamét, ktoré vedú k vzniku dvoch životne dôležitých štruktúr: embrya a endospermu. Po vniknutí peľovej trubice do vajíčka prebehnú dve splynutia súčasne.
Prvé oplodnenie nastáva, keď jedna zo samčích gamét (spermia) zo splývajúcej peľovej trubice splynie s vajíčkovou bunkou (oosférou) v zárodočnom vaku. Tento proces vedie k vytvoreniu diploidnej zygoty (2n), ktorá sa ďalej vyvíja v embryo - základ budúcej rastliny.
Druhé oplodnenie zahŕňa splynutie druhej samčej gaméty s centrálnym jadrom zárodočného vaku, ktoré je však už pred oplodnením diploidné (vzniklo splynutím dvoch polárnych jadier). Splynutím samčej gaméty (n) a diploidného centrálneho jadra (2n) vzniká bunka s triploidným jadrom (3n). Z nej sa sformuje zásobný triploidný endosperm vyživujúci embryo.
Splynutím samčej a samičej gaméty nastalo oplodnenie. Tento mechanizmus je unikátom krytosemenných rastlín, zabezpečujúcim nielen vznik nového jedinca, ale aj jeho ranú výživu. Endosperm je špecializované výživné pletivo, ktoré zásobuje vyvíjajúce sa embryo živinami. Jeho triploidná povaha umožňuje prenos vlastností od oboch rodičov, pričom u niektorých rastlín sa ukazuje, že otcovský genóm hrá významnejšiu úlohu v charakteristikách endospermu. Dvojité oplodnenie teda nie je len mechanizmom vzniku nového jedinca, ale aj zabezpečením jeho výživy počas raného vývinu.
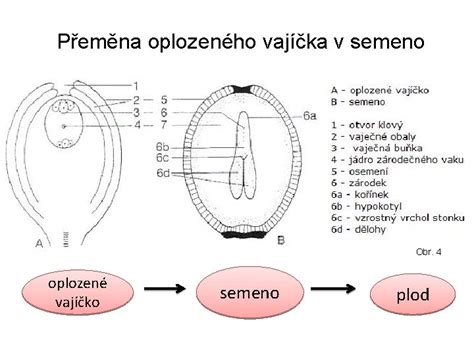
Z post-fertilizačných zmien: Vznik embrya, semena a plodu
Zo zygoty sa ďalším delením a diferenciáciou vyvíja embryo. Proces embryogenézy u krytosemenných rastlín je komplexný. Z oplodneného vajíčka sa vyvíja semeno chránené pevným osemením (testa), ktoré vzniklo z pôvodných vajíčkových obalov - integumentov. Z vnútornej vrstvy integumentu vzniká osemenie. Pre úspešný vývin a následné klíčenie sa v semene, predovšetkým v endosperme, koncentrujú kľúčové živiny a veľké množstvo fosforu. Semeno následne stratí väčšinu vody (jej obsah klesne na 5-20 %) a prechádza do ochranného stavu spánku (dormancie).
Dvojité oplodnenie nie je len mechanizmom vzniku nového jedinca, ale aj zabezpečením jeho výživy počas raného vývinu. Endosperm, ktorý vzniká z triploidného jadra, je špecializované výživné pletivo, ktoré zásobuje vyvíjajúce sa embryo živinami. Antipódy a synergidy, ktoré boli súčasťou zárodočného mieška, po oplodnení zaniknú.
Steny semenníka sa premenia na oplodie (perikarp), ktoré tvorí plod. Z morfologického hľadiska je plod rozmnožovací orgán výlučne krytosemenných rastlín, pretože vzniká zo steny semenníka (alebo celého piestika) po oplodnení vajíčka. Plody a semená slúžia na ochranu a šírenie potomstva.
Evolučné pozadie: Rozmnožovanie a rodozmena v rastlinnej ríši
Rozmnožovanie je základnou funkciou života, zodpovednou za vytváranie nových jedincov a zabezpečenie prežitia druhu. Je neoddeliteľnou súčasťou individuálneho vývinu jedinca, ontogenézy. U živočíchov a rastlín môžeme rozlíšiť dva primárne spôsoby rozmnožovania: nepohlavné (asexuálne, vegetatívne) a pohlavné (sexuálne, generatívne).
Nepohlavné rozmnožovanie (vegetatívne, asexuálne, monogónia) je charakteristické tým, že nový jedinec vzniká priamo z materského organizmu, bez účasti špecializovaných pohlavných buniek. Tento proces prebieha prostredníctvom mitotického delenia somatických buniek. Výsledkom sú geneticky identické potomky, tzv. klony. Hoci tento spôsob umožňuje rýchle šírenie a kolonizáciu prostredia, obmedzuje genetickú variabilitu, čo môže byť nevýhodné pri meniacich sa podmienkach. Jednoduchšie organizmy využívajú bunkové delenie (sinice, jednobunkové riasy), fragmentáciu stielky (spájavky) či tvorbu výtrusov. Vyššie rastliny si na vegetatívne rozmnožovanie vyvinuli špecializované orgány vzniknuté premenou stonky, koreňa či listov, ako sú rozmnožovacie cibuľky (tulipán, cesnak), podzemky (paprade, pýr), podzemkové hľuzy (zemiak), stonkové hľuzy (kaleráb), koreňové hľuzy (reďkovka) alebo poplazy (jahoda). Výnimočne môže semeno vzniknúť aj úplne bez oplodnenia - tento jav produkcie klonálneho semena označujeme ako apomixia.
Pohlavné rozmnožovanie (sexuálne, generatívne, digénne, amfigónia) naopak zahŕňa splynutie dvoch pohlavných buniek, gamét. Tieto gaméty, samičia (makrogaméta, vajíčko, oosféra) a samčia (mikrogaméta, spermia, spermatická bunka), vznikajú redukčným meiotickým delením a sú haploidné (obsahujú polovičný počet chromozómov). Ich splynutím vzniká diploidná zygota, základ nového jedinca s kombinovanou genetickou informáciou od oboch rodičov. Táto genetická variabilita je kľúčová pre adaptáciu druhu na meniace sa prostredie.
Rodozmena (metagenéza) predstavuje životný cyklus rastlín charakterizovaný striedaním dvoch geneticky odlišných generácií: gametofytu a sporofytu. Gametofyt (pohlavná generácia) tvoria bunky s jedným súborom chromozómov (haploidný počet, n). Vzniká mitotickým delením zo spóry a neskôr vytvára pohlavné orgány (gametangiá), ktoré mitózou produkujú pohlavné bunky (gaméty). Plemenníčky (anterídiá) tvoria samčie gaméty (často bičíkaté spermatozoidy vyžadujúce vodu), zatiaľ čo zárodočníky (archegóniá) tvoria samičiu bunku (oosféra). Sporofyt (nepohlavná generácia) disponuje homologickými chromozómami (diploidný počet, 2n) a vzniká mitotickým delením zygoty.
Prechod rastlín na súš znamenal evolučný tlak na ochranu citlivých fáz životného cyklu. Vývoj smeroval k posilneniu diploidného sporofytu a redukcii haploidného gametofytu. Prvé suchozemské rastliny, ako ryniorasty, mali ešte primitívnejšiu rovnakotvarú rodozmenu. Výraznou evolučnou výnimkou sú machorasty (Bryophyta), u ktorých dominuje haploidný gametofyt (zelený mach), pričom sporofyt (stopka s výtrusnicou) je fyziologicky úplne závislý od gametofytu. U cievnatých výtrusných rastlín už dominuje mohutný diploidný sporofyt, zatiaľ čo ich drobný gametofyt (prvorast) je síce zredukovaný, ale žije nezávislým životom.
Pri semenných rastlinách, do ktorých patria aj krytosemenné, je gametofyt extrémne zredukovaný a trvalo ukrytý v materskom sporofyte. Samčí gametofyt krytosemenných rastlín je redukovaný na 2-3 bunky (peľové zrnko) a samičí gametofyt na 7 buniek (zárodočný vak). Toto ukrytie gametofytu v rámci sporofytu predstavuje adaptáciu na suchozemský život a ochranu citlivých reprodukčných štruktúr pred nepriaznivými podmienkami.

Porovnanie s nahosemennými rastlinami: Kontrastné stratégie rozmnožovania
Nahosemenné rastliny (Gymnospermae) tvoria evolučný prechod medzi výtrusnými cievnatými a krytosemennými rastlinami. Vyvinuli sa koncom starších prvohôr a ich najdôležitejším znakom je tvorba semien. Avšak, vajíčka a z nich vzniknuté semená nie sú ukryté v semenníku a netvoria plody, ale ležia voľne exponované na semenných šupinách, sú „nahé“. Chýbajú im typické kvety s obalmi a bliznou. Kvety sa zoskupujú do jednopohlavných samčích a samičích šišiek (strobilov). Tento fakt treba zdôrazniť, pretože používanie termínu „plod“ v súvislosti s nahosemennými rastlinami je laická a botanicky nepresná terminológia. Keďže nemajú semenník, po oplodnení vajíčka z neho vznikne len nahé semeno bez oplodia, a teda z definície nemôže vzniknúť plod.
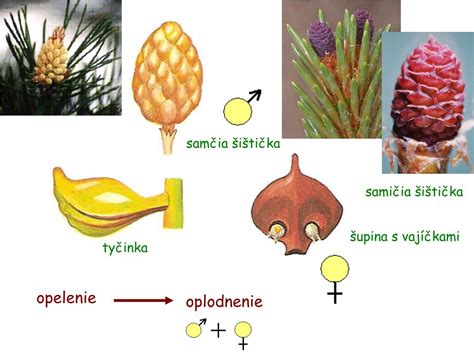
Rozmnožovanie nahosemenných rastlín sa líši od krytosemenných v niekoľkých kľúčových aspektoch. Vezmime si ako príklad borovicu lesnú (Pinus sylvestris). Na borovici, ktorá je sporofytom, vyrastajú samčie a samičie šištice (strobily). Samičie šištice (megastrobily) vyrastajú na vrchných konároch a sú tvorené zdrevnateným vretenom, na ktorom sú špirálovito usporiadané podporné šupiny a v ich úžľabí vyrastajú semenné šupiny. U borovice sú podporné šupiny zakrpatené a semennými šupinami prerastené natoľko, že sú morfologicky takmer nerozoznateľné. Na vrchnej strane semenných šupín sú pri báze lokalizované dve vajíčka.
Vajíčka nahosemenných rastlín sú krasinucelátne. Nucelus je pokrytý výraznou pokožkou a obklopený veľkým počtom parenchymatických buniek. Na vrchole je nucelus sformovaný do peľovej komory, ktorá vzniká buď lýzou skupiny buniek v apikálnej časti alebo nápadným naddvihnutím. Oba tieto útvary slúžia na vytvorenie vhodných podmienok pre zachytenie a vyklíčenie peľového zrna. Integument, ktorý je u nahosemenných rastlín obvykle len jeden, postupne prerastá nucelus a vytvorí sa mikropyla, ktorá podobne ako blizna krytosemenných rastlín produkuje polinačnú kvapku.
Z materskej bunky megaspóry sa redukčným delením vzniká tetráda haploidných megaspór. Pri Pinus sylvestris v procese heterotypického delenia vznikajú dve nerovnako veľké bunky, pričom sa ďalej vyvíja len bazálna megaspóra. Z jej niekoľkonásobného mitotického delenia vzniká voľnojadrové prothálium. Výskyt veľkého počtu voľných jadier je charakteristickým znakom všetkých nahosemenných rastlín. Následne sa vytvoria bunkové steny a z týchto haploidných buniek vzniká endosperm a archegóniá. Endosperm nahosemenných rastlín je haploidný a má charakter zásobného parenchýmu, obsahujúceho predovšetkým škrob, aleuronové zrná a lipidy. U Pinus sylvestris samičí gametofyt pozostáva z haploidného endospermu a z dvoch archegónií umiestnených v oblasti mikropyly, pričom v každom archegóniu je jedna oosféra. Archegónium je samičí pohlavný orgán rastlín, oosféra je samičia gaméta.
Samčie šištice (mikrostrobily) vyrastajú v spodných etážach stromu, sú tvorené krátkym vretenom a blanitými, špirálovito usporiadanými mikrosporofylmi (tyčinkami) s dvoma alebo niekoľkými mikrosporangiami. Vo vnútri peľového zrna sa v jednom mieste oddelí niekoľko drobných buniek, ktoré predstavujú samčie prothálium a väčšinou po určitom čase zanikajú. Vnútro peľového zrna vypĺňa veľká bunka, ktorá sa mitoticky delí na väčšiu vegetatívnu bunku a menšiu generatívnu bunku. Generatívna bunka sa opäť delí na nástennú a spermatogénnu bunku, z ktorej pri prerastaní peľového vrecúška vzniknú dve spermatické bunky. U primitívnejších skupín nahosemenných rastlín (napr. cykasovitých) sa samčie gaméty vyvíjajú ako obrvené spermatozoidy, zatiaľ čo u ihličnanov sú neobrvené.
Opelenie nahosemenných rastlín je prevažne vetrom. Počas prvej sezóny sa šupiny samičích šištíc so zrelými vajíčkami oddialia od vretena. Peľové zrná dopadajú medzi šupiny a na povrchu vajíčka sa zachytávajú do polinačnej kvapky, ktorá je vylučovaná v oblasti mikropyly. Pri vysýchaní kvapky sa peľové zrno dostane na povrch nucela. Peľové zrno klíči, vzniká peľová trubica (peľové vrecúško), ktoré začne prerastať cez nucelus. Šupiny samičích šištíc sa v tomto období primknú k vretenu. Spermatogénna bunka sa v peľovej trubici rozdelí na dve spermatické bunky. Jedna spermatická bunka oplodní oosféru a vzniká zygota. Ostatné bunky peľovej trubice zanikajú. Na rozdiel od krytosemenných rastlín, u nahosemenných rastlín splynie spermatické jadro s vajcovou bunkou a vytvorí sa zygóta - ide teda o jednoduché oplodnenie, nie dvojité. Zásobné pletivo (endosperm) je haploidné a je vytvorené pred oplodnením.
Vo vývine embrya nahosemenných rastlín sú určité rozdiely oproti krytosemenným. Pre proces embryogenézy je charakteristický jav voľnojadrového delenia, to znamená, že v počiatočných fázach sa jadro zygoty delí, ale nevznikajú bunkové steny. Počet voľnojadrových delení je u jednotlivých rodov rôzny a až potom nastáva celularizácia. Napríklad u Ginkgo biloba nastáva cytokinéza až po vytvorení 256 jadier, u cykasu Dioon edule 1000 a u ihličnanov po vytvorení 2 až 64 jadier. Ďalšou zvláštnosťou ihličnatých rastlín je polyembryónia, pri ktorej vzniká viacero embryí z jedného vajíčka. Napríklad v rode Pinus v prvej fáze voľnojadrového delenia vzniknú štyri jadrá, ktoré putujú k mikropyle a usporiadajú sa do jednej vrstvy, z ktorých sa postupne vyvíjajú embryá. Z oplodneného vajíčka sa vyvíja embryo a celé vajíčko sa premieňa na semeno. Z vnútornej vrstvy integumentu vzniká osemenie a z vonkajšej vrstvy sa diferencuje krídielko, ktoré napomáha rozširovaniu semien vetrom.
Niektoré ihličnany nemajú drevnaté šištice a nie sú ani jednodomé. Napríklad tis obyčajný (Taxus baccata) je dvojdomá drevina, produkuje vajíčka na koncoch krátkych axilárnych výhonkov v mäsitých pohárikovitých mieškoch (arilusoch), ktoré chránia semeno a obľubujú ich vtáky. Tisom chýbajú živicové kanáliky v dreve. Cykasorasty (Cycadophyta), „žijúce fosílie“, sú striktne dvojdomé a na rozdiel od iných nahosemenných rastlín ich opeľuje hmyz (najmä chrobáky). Vytvárajú semená s nápadným mäsitým obalom (sarkotestou), ktoré vzhľadom pripomínajú kôstkovicu. Ginkorasty (Ginkgophyta) predstavujú jedinečnú a starobylú vývojovú líniu, pričom dnes prežíva už len jediný reliktný druh, ginko dvojlaločné (Ginkgo biloba). Je to dvojdomá rastlina, ktorá netvorí klasické šišky, ale mäsité semená s tvrdou kôstkou, ktoré po dozretí silne zapáchajú. Lianovcorasty (Gnetophyta) sú evolučne najpokročilejšou skupinou a dôležitým prechodom ku krytosemenným rastlinám. Ako jediné nahosemenné rastliny už majú v dreve pravé cievy a u niektorých dochádza k dvojitému oplodneniu, čo je zriedkavá konvergentná evolúcia s krytosemennými rastlinami. Extrémnym a fascinujúcim zástupcom je velvíčia podivná (Welwitschia mirabilis) z púšte Namíb.
Význam a úspech krytosemenných rastlín na Zemi
Dvojité oplodnenie predstavuje sofistikovaný mechanizmus, ktorý významne prispel k evolučnému úspechu krytosemenných rastlín, tvoriacich dnes dominantnú skupinu rastlinnej ríše. Zabezpečenie výživy embrya prostredníctvom triploidného endospermu zvyšuje šance na jeho prežitie a úspešné vyklíčenie. Taktiež umožňuje rastlinám efektívnejšie hospodáriť so zdrojmi, keďže výživné pletivo sa tvorí až po úspešnom oplodnení. Na rozdiel od haploidného endospermu nahosemenných rastlín, ktorý sa vyvíja pred oplodnením bez ohľadu na jeho úspešnosť, triploidný endosperm krytosemenných rastlín je energeticky efektívnejší, pretože je výsledkom úspešnej fúzie.
Kryštalizácia týchto výhod, spolu s vývojom kvetov ako lákadiel pre opeľovačov a plodov ako účinných mechanizmov šírenia semien a ich ochrany, umožnila krytosemenným rastlinám kolonizovať takmer všetky ekosystémy na Zemi. Stali sa základom potravy pre mnohé živočíchy, vrátane človeka, a tvoria štrukturálny základ väčšiny suchozemských ekosystémov. Ich morfologická diverzita, adaptabilita a efektívne reprodukčné stratégie, z ktorých dvojité oplodnenie je ústredné, sú kľúčom k ich bezkonkurenčnej dominancii.
Proces opeľovania a oplodnenia 🌷| Postup krok za krokom
tags: #oplodnenie #krytosemennych #rastlin